
Changes in Macrozoobenthos Community after Aquatic Plant Restoration in the Northern Venice Lagoon (IT)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
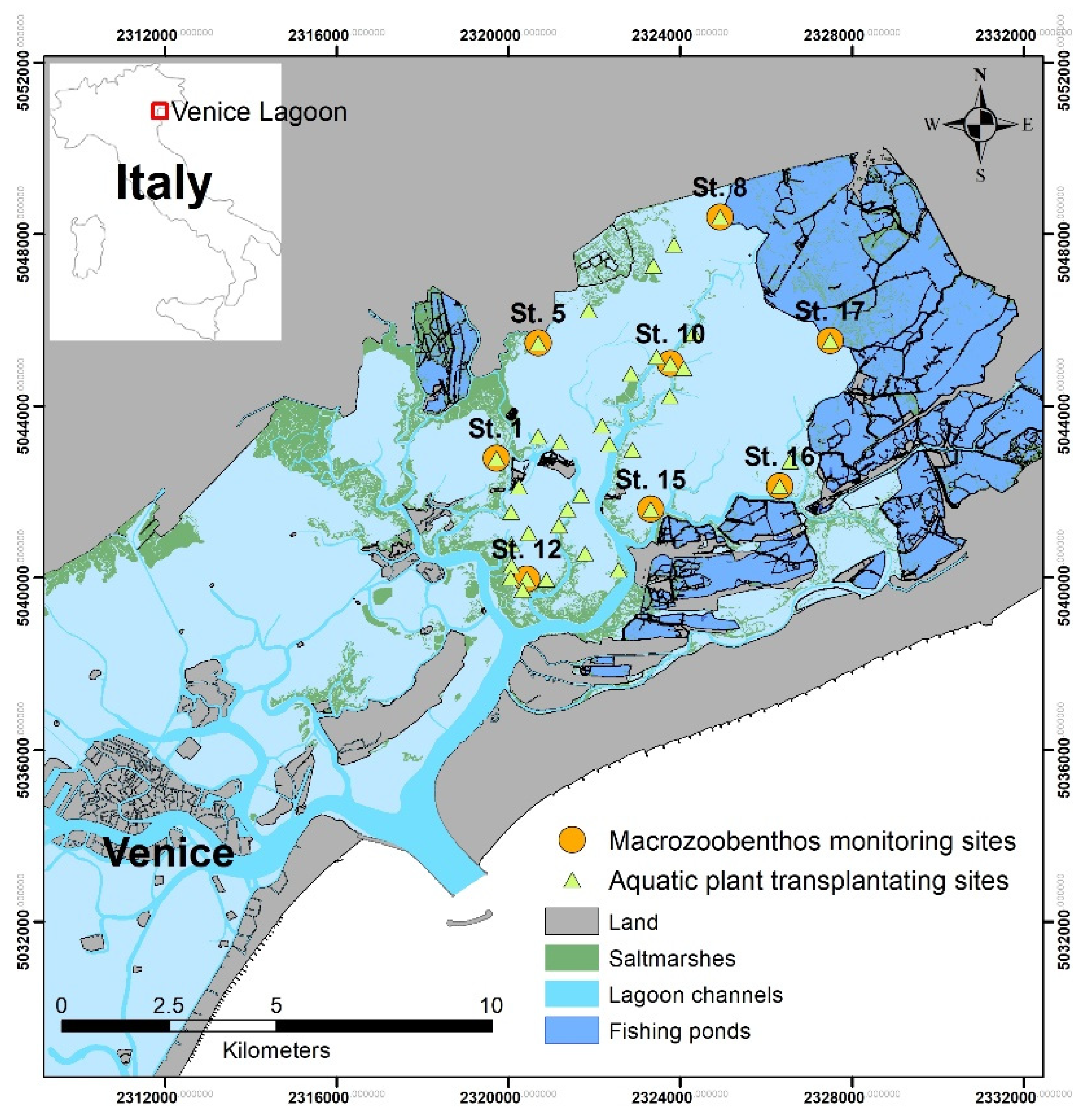
2.1. Study Area and Sampling Site
2.2. Sampling Procedure and Sample Treatment
2.3. Data Analysis
3. Results
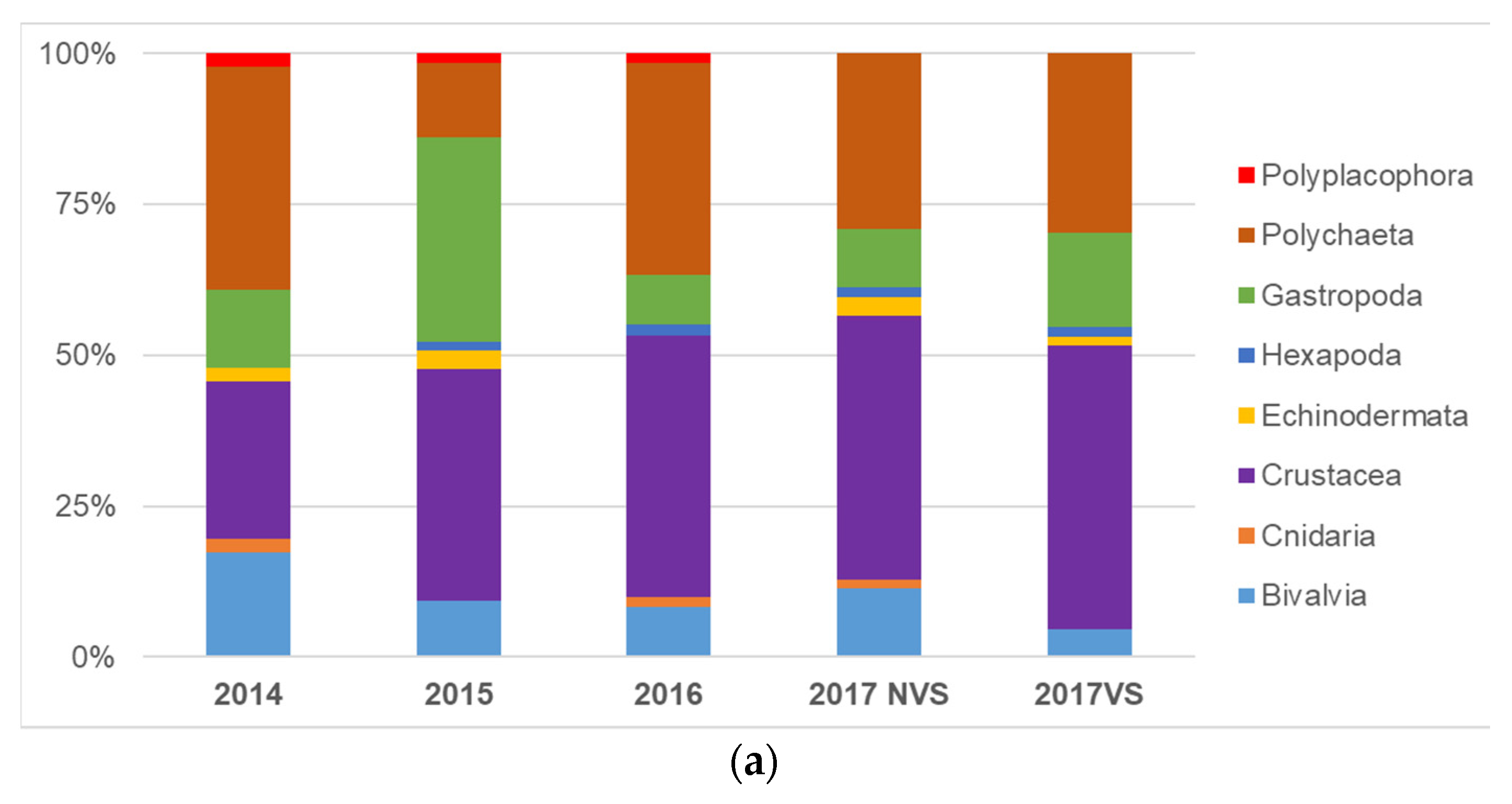
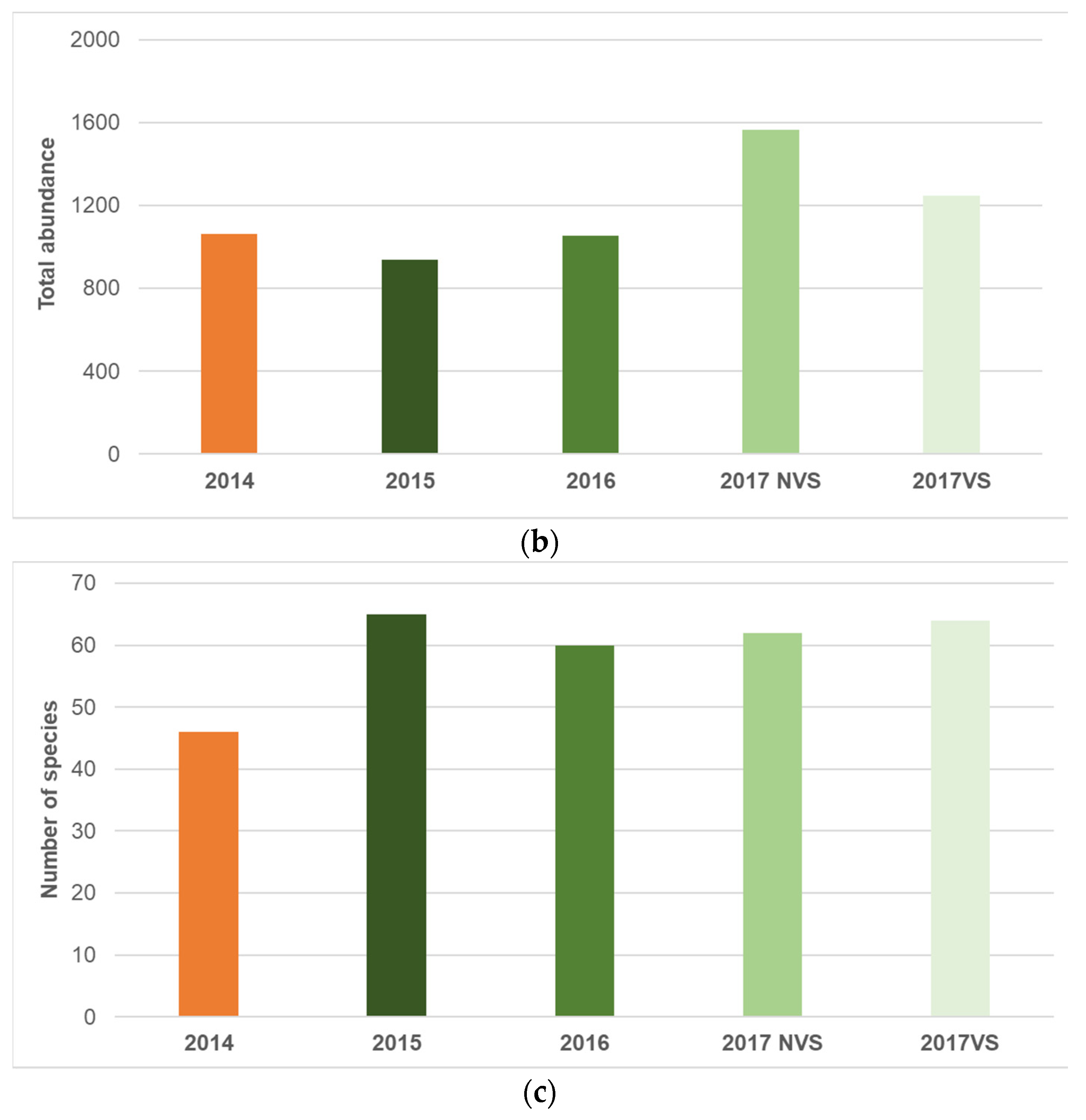
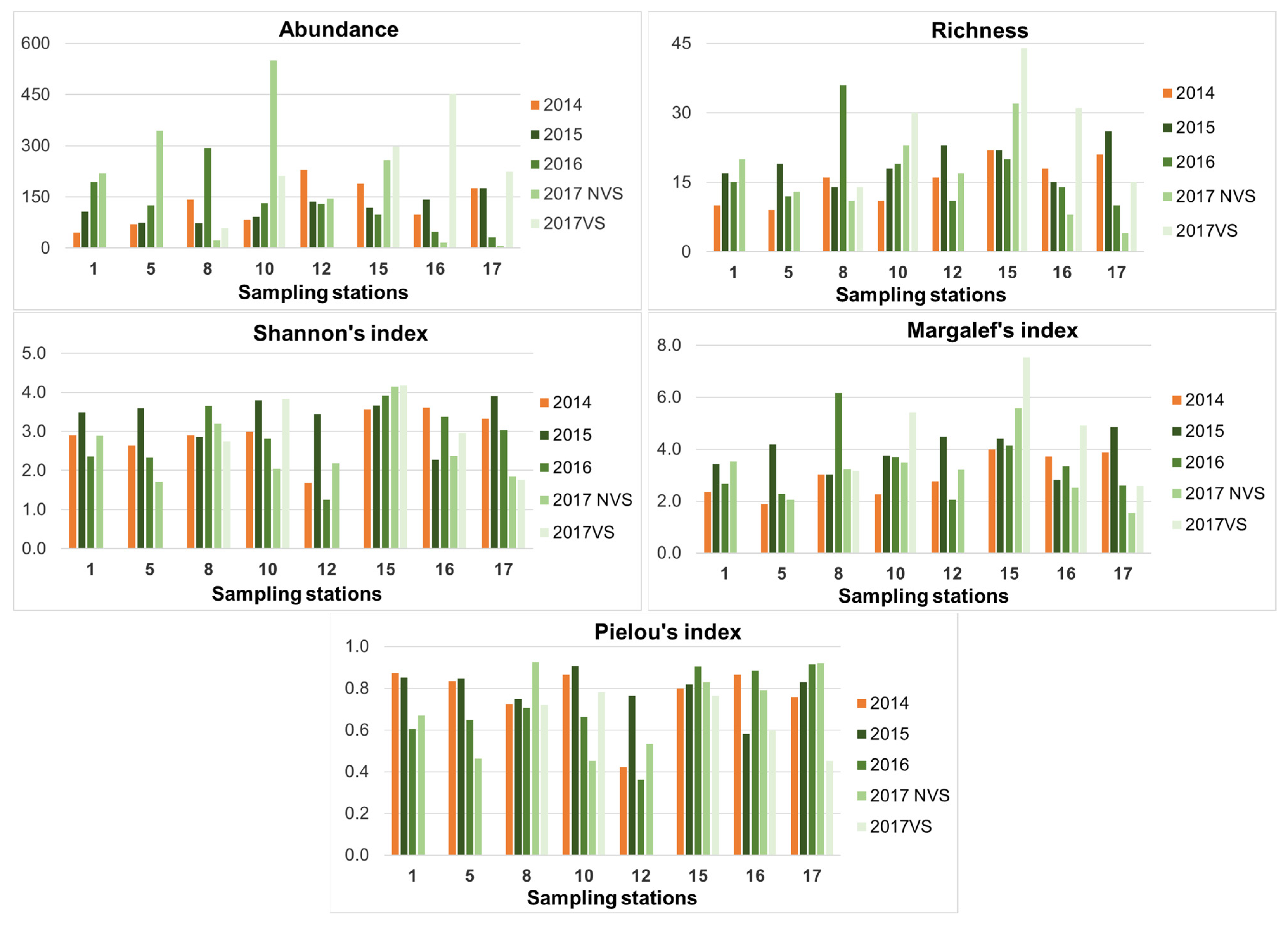
3.1. Univariate Analysis
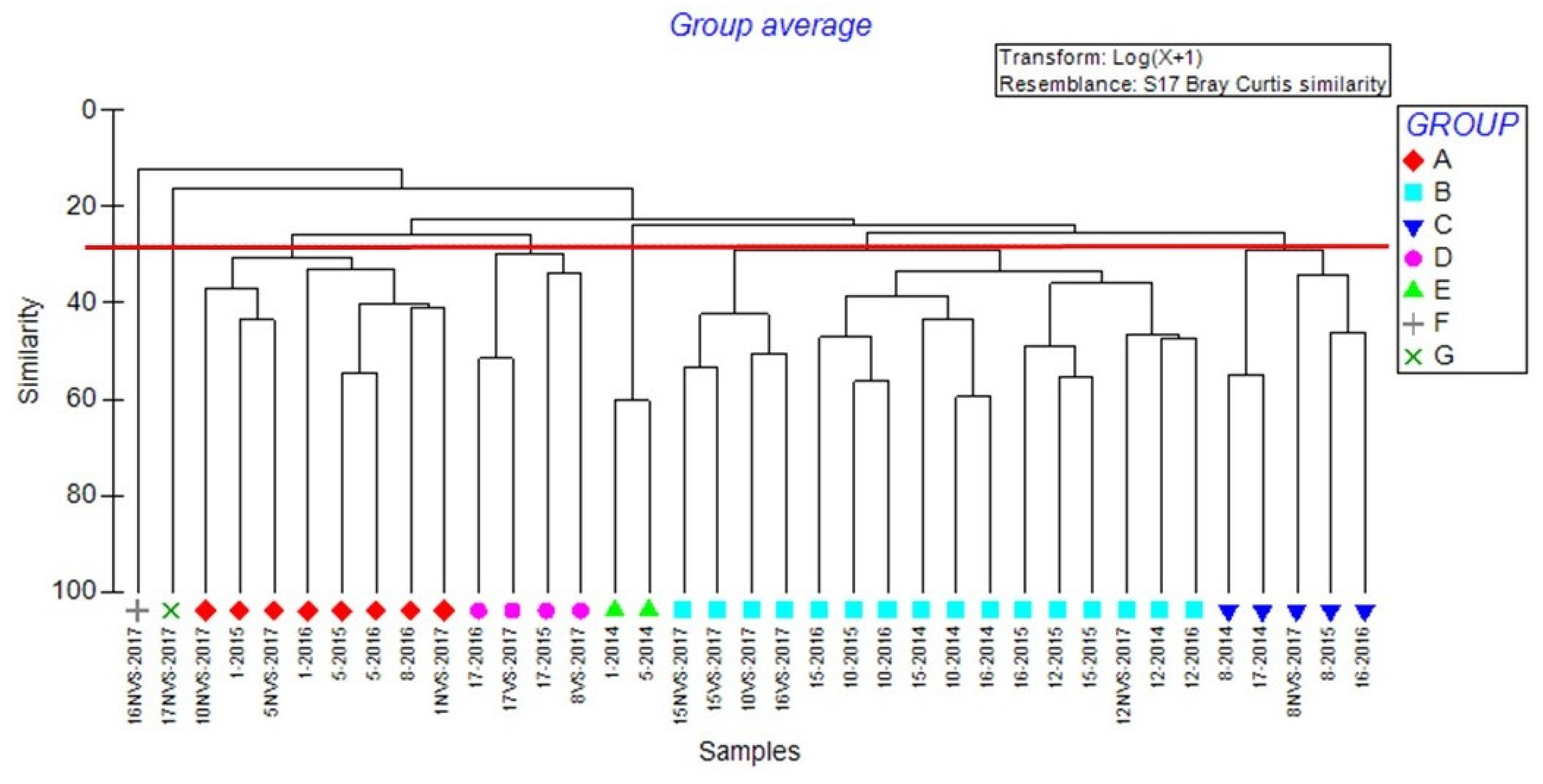
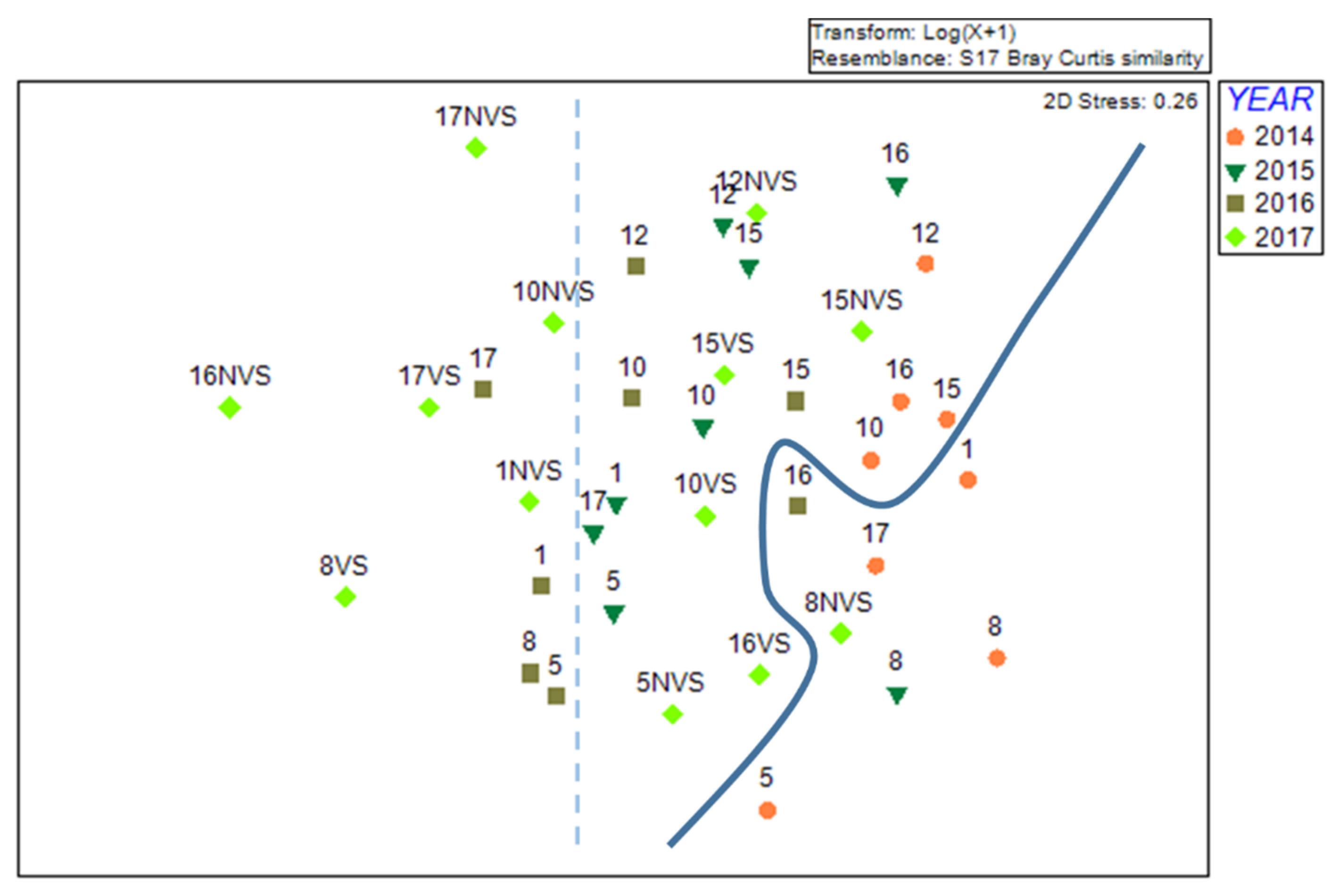
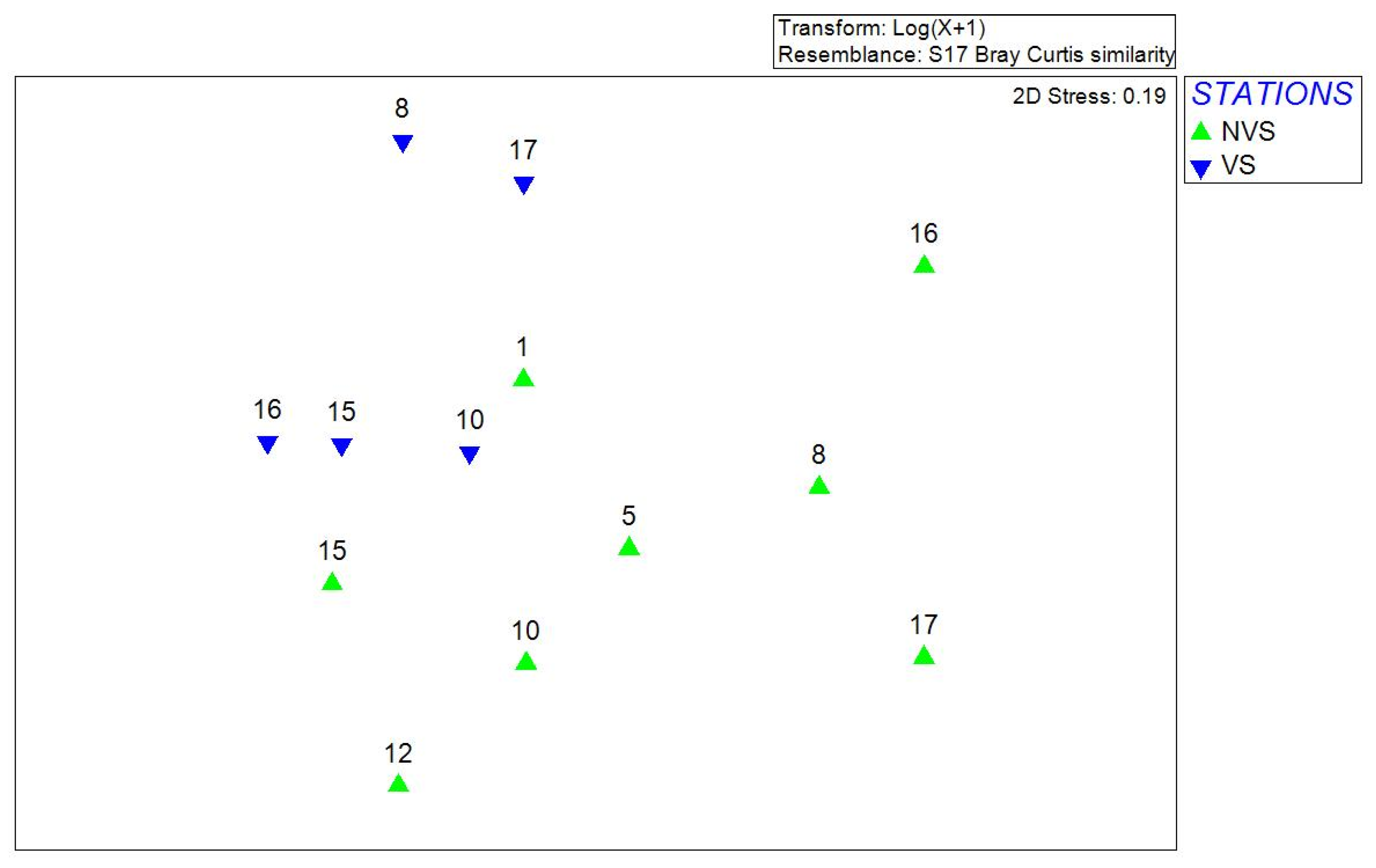
3.2. Multivariate Analysis
3.3. Ecological Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borja, A.; Franco, J.; Pérez, V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and Coastal environments. Mar. Poll. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M. Benthic fauna: Collection and identification of macrobenthic invertebrates. Terre Environ. 2010, 88, 253–261. [Google Scholar]
- Snelgrove, P.V.R. The biodiversity of macrofaunal organisms in marine sediments. Biodivers. Conserv. 1998, 7, 1123–1132. [Google Scholar] [CrossRef]
- Ysebaert, T.; Meire, P.; Herman, P.M.J.; Verbeek, H. Macrobenthic species response surfaces along estuarine gradients: Prediction by logistic regression. Mar. Ecol. Prog. Ser. 2002, 225, 79–85. [Google Scholar] [CrossRef]
- Dauer, D.M. Biological criteria, environmental health and estuarine macrobenthic community structure. Mar. Poll. Bull. 1993, 26, 249–257. [Google Scholar] [CrossRef]
- López-López, E.; Sedeño-Díaz, J.E. Biological indicators of water quality: The role of fish and macroinvertebrates as indicators of water quality. In Environmental Indicators; Armon, R.H., Hänninen, O., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 643–661. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Volpi Ghirardini, A. A review of terms and definitions to categorise estuaries, lagoons and associated environments. Mar. Freshw. Res. 2009, 60, 497–509. [Google Scholar] [CrossRef]
- Facca, C.; Pellegrino, N.; Ceoldo, S.; Tibaldo, M.; Sfriso, A. Trophic condition in the waters of the Venice Lagoon (Northern Adriatic Sea, Italy). Open Oceanogr. J. 2011, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dauvin, J.C. Paradox of estuarine quality: Benthic indicators and indices, consensus or debate for the future. Mar. Poll. Bull. 2007, 55, 271–281. [Google Scholar] [CrossRef]
- Mattila, J.; Chaplin, G.; Eilers, M.R.; Heck, K.L.; O’Neal, J.P.; Valentine, J.F. Spatial and diurnal distribution of invertebrate and fish fauna of a Zostera marina bed and nearby unvegetated sediments in Damariscotta River, Maine (USA). J. Sea Res. 1999, 41, 321–332. [Google Scholar] [CrossRef]
- Bowden, D.A.; Rowden, A.A.; Martin, J.A. Effect of patch size and in-patch location on the infaunal macroinvertebrate assemblages of Zostera marina seagrass beds. J. Exp. Mar. Biol. Ecol. 2001, 259, 133–154. [Google Scholar] [CrossRef]
- Sfriso, A.; Birkemeyer, T.; Ghetti, P.F. Benthic macrofauna changes in areas of Venice lagoon populated by seagrasses or seaweeds. Mar. Environ. Res. 2001, 52, 323–349. [Google Scholar] [CrossRef]
- Fredriksen, S.; De Backer, A.; Böstrom, C.; Christie, H. Infauna from Zostera marina L. meadows in Norway. Differences in vegetated and unvegetated areas. Mar. Biol. Res. 2010, 6, 189–200. [Google Scholar] [CrossRef]
- Barnes, R.S.K.; Barnes, M.K.S. Shore height and differentials between macrobenthic assemblages in vegetated and unvegetated areas of an intertidal sandflat. Estuar. Coast. Shelf Sci. 2012, 106, 112–120. [Google Scholar] [CrossRef]
- Orth, R.J.; Heck, K.L., Jr.; van Montfrans, J. Faunal communities in seagrass beds: A review of the influence of plant structure and prey characteristics on predator-prey relationships. Estuaries 1984, 7, 339–350. [Google Scholar] [CrossRef]
- Mazzella, L.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Plant-animal trophic relationships in the Posidonia oceanica ecosystem of the Mediterranean Sea: A review. In Plant–Animals Interactions in the Marine Benthos. Systematics Association; John, D.M., Hawkins, S.J., Price, J.H., Eds.; Clarendon Press: Oxford, UK, 1992; Special Volume 46, pp. 165–187. [Google Scholar]
- Duffy, J.E. Biodiversity and the functioning of seagrass ecosystems. Mar. Ecol. Progr. Ser. 2006, 311, 233–250. [Google Scholar] [CrossRef] [Green Version]
- Cossu, R.; De Fraja Frangipane, E.; Degobbis, D.; Orio, A.A.; Andreottola, G. Pollution and eutrophication in the Venice Lagoon. Water Sci. Technol. 1987, 19, 813–822. [Google Scholar] [CrossRef]
- Sfriso, A.; Pavoni, B.; Marcomini, A.; Orio, A.A. Macroalgae, Nutrient Cycles, and Polluntants in the Lagoon of Venice. CERF 1992, 15, 517–528. [Google Scholar] [CrossRef]
- Solidoro, C.; Bandelj, V.; Bernardi, F.A.; Camatti, E.; Ciavatta, S.; Cossarini, G.; Facca, C.; Franzoi, P.; Libralato, S.; Melaku Canu, D.; et al. Response of the Venice Lagoon eco system to natural and anthropogenic pressures over the last 50 years. In Coastal Lagoons. Critical Habitats of Environmental Change; Kennish, M.J., Pearl, H.W., Eds.; CRC Press: Boca Raton, FL, USA, 2010; Chapter 19; pp. 483–511. [Google Scholar] [CrossRef]
- Solidoro, C.; Melaku Canu, D.; Cucco, A.; Umgiesser, G. A partition of the Venice lagoon based of physical properties and analysis of general circulation. J. Mar. Syst. 2004, 51, 147–160. [Google Scholar] [CrossRef]
- Sorokin, Y.I.; Sorokin, P.Y.; Giovanardi, O.; Dalla Venezia, L. Study of the ecosystem of the lagoon of Venice, with emphasis on anthropogenic impact. Mar. Ecol. Prog. Ser. 1996, 141, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; Facca, C. Distribution and production of macrophytes in the lagoon of Venice. Comparison of actual and past abundance. Hydrobiologia 2007, 577, 71–85. [Google Scholar] [CrossRef]
- Facca, C.; Ceoldo, S.; Pellegrino, N.; Sfriso, A. Natural recovery and planned intervention in coastal wetlands: Venice Lagoon (Northern Adriatic Sea, Italy) as a case study. Sci. World J. 2014, 968618. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, A.; Buosi, A.; Facca, C.; Sfriso, A.A.; Tomio, Y.; Juhmani, A.-S.; Wolf, M.A.; Franzoi, P.; Scapin, L.; Ponis, E.; et al. Environmental restoration by aquatic angiosperm transplants in transitional water systems: The Venice Lagoon as a case study. Sci. Total Environ. 2021, 795, 148859. [Google Scholar] [CrossRef] [PubMed]
- Lefcheck, J.S.; Orth, R.J.; Dennison, W.C.; Wilcox, D.J.; Murphy, R.R.; Keisman, J.; Gurbisz, C.; Hannam, M.; Landry, J.B.; Moore, K.A.; et al. Long-term nutrient reductions lead to the unprecedented recovery of a temperate coastal region. Proc. Natl. Acad. Sci. USA 2018, 115, 3658–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curiel, D.; Miotti, C.; Dri, C.; Gentilin, S.; Checchin, E.; Rismondo, A.; Caniglia, G. Dinamiche di crescita ed epifitismo delle fanerogame marine della Laguna di Venezia. Lav. Soc. Ven. Sc. Nat 2006, 31, 67–76. [Google Scholar]
- Rismondo, A.; Curiel, D.; Scarton, F.; Mion, D.; Caniglia, G. A new seagrass map for the Venice Lagoon. In Proceedings of the Sixth International Conference on the Mediterranean Coastal Environment, Ravenna, Italy, 7–11 October 2003; Middle East Technical University Publications: Ankara, Turkey, 2003; pp. 843–852. [Google Scholar]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Validation of the Macrophyte Quality Index (MaQI) set up to assess the ecological status of Italian marine transitional environments. Hydrobiologia 2009, 617, 117–141. [Google Scholar] [CrossRef]
- Costanza, R.; D’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Cacciatore, F.; Bonometto, A.; Paganini, E.; Sfriso, A.; Novello, M.; Parati, P.; Gabellini, M.; Boscolo Brusà, R. Balance between the Reliability of Classification and Sampling Effort: A Multi-Approach for the Water Framework Directive (WFD) Ecological Status Applied to the Venice Lagoon (Italy). Water 2019, 11, 1572. [Google Scholar] [CrossRef] [Green Version]
- Tagliapietra, D.; Pavan, M.; Wagner, C. Macrobenthic Community Changes Related to Eutrophication in Palude della Rosa (Venetian Lagoon, Italy). Estuar. Coast. Shelf Sci. 1998, 47, 217–226. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Pavan, M.; Wagner, C. Benthic patterns in a salt marsh basin: A snapshot of Palude della Rosa (Venetian Lagoon, Italy). Wetl. Ecol. Manag. 2000, 8, 287–292. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer Version 6.0; PRIMER-e: Plymouth, UK, 2006. [Google Scholar]
- StatSoft, Inc. STATISTICA. Data Analysis Software System, Version 6; StatSoft, Inc.: Tulsa, OK, USA, 2001.
- Muxika, I.; Borja, A.; Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 16–29. [Google Scholar] [CrossRef]
- Pranovi, F.; Da Ponte, F.; Torricelli, P. Application of biotic indices and relationship with structural and functional features of macrobenthic community in the lagoon of Venice: An example over a long time series of data. Mar. Poll. Bull. 2007, 54, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Sigovini, M. Multiscale Dynamics of Zoobenthic Communities and Relationships with Environmental Factors in the Lagoon of Venice. Ph.D. Thesis, Venice Ca’ Foscari University, Venice, Italy, 2010. [Google Scholar]
- Mistri, M.; Fano, E.; Rossi, G.; Caselli, K.; Rossi, R. Variability in macrobenthos communities in the Valli di Comacchio, Northern Italy, a hypereutrophized lagoonal ecosystem. Estuar. Coast. Shelf Sci. 2000, 51, 599–611. [Google Scholar] [CrossRef]
- Marzano, C.N.; Liaci, L.S.; Fianchini, A.; Gravina, F.; Mercurio, M.; Corriero, G. Distribution, persistence and change in the macrobenthos of the lagoon of Lesina (Apulia, southern Adriatic Sea). Oceanol. Acta 2003, 26, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Pranovi, F.; Curiel, D.; Rismondo, A.; Marzocchi, M.; Scattolin, M. Variations of the macrobenthic community in a seagrass transplanted area of the Lagoon of Venice. Sci. Mar. 2000, 64, 303–310. [Google Scholar] [CrossRef]
- Da Ros, Z.; Corinaldesi, C.; Dell’Anno, A.; Gambi, C.; Torsani, F.; Danovaro, R. Restoration of Cymodocea nodosa seagrass meadows: Efficiency and ecological implications. Restor. Ecol. 2021, 29, e13313. [Google Scholar] [CrossRef]
- Ambo-Rappe, R.; Rani, C. Physical structure of artificial seagrass affects macrozoobenthic community recruitment. J. Phys. Conf. Ser. 2018, 979, 012006. [Google Scholar] [CrossRef] [Green Version]
- Asriani, N.; Ambo-Rappe, R.; Lanuru, M.; Williams, S.L. Macrozoobenthos community structure in restored seagrass, natural seagrass and seagrassless areas around Badi Island, Indonesia. IOP Conf. Ser. Earth Environ. 2019, 253, 012034. [Google Scholar] [CrossRef]
- Homziak, J.; Fonseca, M.S.; Kenworthy, W.J. Macrobenthic community structure in a transplanted eelgrass (Zostera marina) meadow. Mar. Ecol. Prog. Ser. 1982, 9, 211–221. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Meyer, D.L.; Hall, M.O. Development of planted seagrass beds in Tampa Bay, Florida, USA. II. Faunal components. Mar. Ecol. Prog. Ser. 1996, 132, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Posey, M.H. Community changes associated with the spread of an introduced seagrass, Zostera japonica. Ecology 1988, 69, 974–983. [Google Scholar] [CrossRef]
- Lewis, F.G. Distribution of macrobenthic crustacean associated with Thalassia, Halodule and bare sand substrata. Mar. Ecol. Prog. Ser. 1984, 19, 101–113. [Google Scholar] [CrossRef]
- Valdez, S.R.; Shaver, E.C.; Keller, D.A.; Morton, J.P.; Zhang, Y.S.; Wiernicki, C.; Chen, C.; Martinez, C.; Silliman, B.R. A survey of benthic invertebrate communities in native and non-native seagrass beds in St. John, USVI. Aquat. Bot. 2021, 175, 103448. [Google Scholar] [CrossRef]
- Sheridan, P.; Henderson, C.; McMahan, G. Fauna of natural seagrass and transplanted Halodule wrightii (shoal-grass) beds in Galveston Bay, Texas. Restor. Ecol. 2003, 11, 139–154. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Kenworthy, W.J.; Thayer, G.W. Guidelines for the Conservation and Restoration of Seagrasses in the United States and Adjacent Waters; National Oceanic and Atmospheric Administration, Coastal Ocean Office: Charleston, SC, USA, 1998; Volume 55.
- Sfriso, A.; Marcomini, A. Macrophyte production in a shallow coastal lagoon. Part I: Coupling with chemico-physical parameters and nutrient concentrations in waters. Mar. Environ. Res. 1997, 44, 351–375. [Google Scholar] [CrossRef]
- Sfriso, A.; Marcomini, A. Macrophyte production in a shallow coastal lagoon. Part II: Coupling with sediment, SPM and tissue carbon, nitrogen and phosphorus concentrations. Mar. Environ. Res. 1999, 47, 285–309. [Google Scholar] [CrossRef]
- Hauxwell, J.; Valiela, I. Effects of nutrient loading on shallow seagrass-dominated coastal systems: Patterns and processes. In Estuarine Nutrient Cycling: The Influence of Primary Producers; Springer: Dordrecht, The Netherlands, 2004; pp. 59–92. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Pessa, G.; Cornello, M.; Zitelli, A.; Magni, P. Temporal distribution of intertidal macrozoobenthic assemblages in a Nanozostera noltii-dominated area (Lagoon of Venice). Mar. Environ. Res. 2016, 114, 31–39. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Tomio, Y.; Juhmani, A.-S.; Facca, C.; Sfriso, A.A.; Franzoi, P.; Scapin, L.; Bonometto, A.; Ponis, E.; et al. Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems. Water 2019, 11, 2135. [Google Scholar] [CrossRef] [Green Version]
- Guerzoni, S.; Tagliapietra, D. Atlante Della Laguna: Venezia Tra Terra e Mare; Marsilio: Venice, Italy, 2006. [Google Scholar]
- Maggiore, F.; Keppel, E. Biodiversity and distribution of polychaetes and molluscs along the Dese estuary (Lagoon of Venice, Italy). Hydrobiologia 2007, 588, 189–203. [Google Scholar] [CrossRef]
- Turner, S.J.; Kendall, M.A.A. comparison of vegetated and unvegetated soft-sediment macrobenthic communities in the River Yealm, south-western Britain. J. Mar. Biolog. Assoc. UK 1999, 79, 741–743. [Google Scholar] [CrossRef]
- Qiu, D.; Yan, J.; Ma, X.; Gao, F.; Wang, F.; Wen, L.; Bai, J.; Cui, B. How vegetation influence the macrobenthos distribution in different saltmarsh zones along coastal topographic gradients. Mar. Environ. Res. 2019, 151, 104767. [Google Scholar] [CrossRef]
- Sconfietti, R. Researches on spatial distribution of amphipods, isopods and tanaids (Peracarida) in a Mediterranean estuary (River Dese, Lagoon of Venice). Crustaceana 1988, 55, 193–201. [Google Scholar] [CrossRef]
- Patarnello, T.; Bisol, P.M.; Battaglia, B. Studies on differential fitness of PGI genotypes with regard to temperature in Gammarus insensibilis (Crustacea: Amphipoda). Mar. Biol. 1989, 102, 355–359. [Google Scholar] [CrossRef]
- Conradi, M.; Lopez-Gonzalez, P.J.; Garcia-Gomez, J.C. The amphipod community as a bioindicator in Algeciras Bay (Southern Iberian Peninsula) based on a spatio-temporal distribution. Mar. Ecol. 1997, 18, 97–111. [Google Scholar] [CrossRef]
- Zaabar, W.; Zakhama-Sraieb, R.; Charfi-Cheikhrouha, F.; Achouri, M.S. Abundance and diversity of amphipods (Crustacea: Peracarida) on shallow algae and seagrass in lagoonal ecosystem of the Mediterranean Tunisian coast. Zool. Stud. 2015, 54, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, E.; Anadon, R.; Fernández, C. Life histories and growth of the gastropods Bittium reticulatum and Barleeia unifasciata inhabiting the seaweed Gelidium latifolium. J. Molluscan Stud. 1988, 54, 119–129. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Silvestri, S.; Ghetti, P.F. Role of macroalgal biomass and clam fishing on spatial and temporal changes in N and P sedimentary pools in the central part of the Venice lagoon. Oceanol. Acta 2003, 26, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.B.; Haderlie, E.C. The distribution of Nephtys cirrosa and N. hombergi on the south-western coasts of England and Wales. J. Anim. Ecol. 1960, 29, 117–147. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. 1979, 17, 193–284. [Google Scholar]
- Pitacco, V.; Orlando-Bonaca, M.; Mavrič, B.; Popović, A.; Lipej, L. Mollusc fauna associated with the Cystoseira algal associations in the Gulf of Trieste (Northern Adriatic Sea). Mediterr. Mar. Sci. 2014, 15, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Dauer, D.; Elliott, M.; Simenstad, C. Medium- and long-term recovery of estuarine and coastal ecosystems: Patterns, rates and restoration effectiveness. Estuar. Coasts 2010, 33, 1249–1260. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | |||||
|---|---|---|---|---|---|
| Stations | 2014 | 2015 | 2016 | 2017 NVS | 2017 VS |
| 1 | 0.73 | 0.91 | 0.7 | 0.91 | |
| 5 | 0.69 | 0.96 | 0.65 | 0.58 | |
| 8 | 0.53 | 0.51 | 0.8 | 0.59 | 0.59 |
| 10 | 0.54 | 0.68 | 0.66 | 0.6 | 0.8 |
| 12 | 0.68 | 0.95 | 0.61 | 0.79 | |
| 15 | 0.59 | 0.69 | 0.68 | 0.76 | 0.88 |
| 16 | 0.62 | 0.46 | 0.55 | 0.44 | 0.78 |
| 17 | 0.67 | 0.76 | 0.55 | 0.49 | 0.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oselladore, F.; Bernarello, V.; Cacciatore, F.; Cornello, M.; Boscolo Brusà, R.; Sfriso, A.; Bonometto, A. Changes in Macrozoobenthos Community after Aquatic Plant Restoration in the Northern Venice Lagoon (IT). Int. J. Environ. Res. Public Health 2022, 19, 4838. https://doi.org/10.3390/ijerph19084838
Oselladore F, Bernarello V, Cacciatore F, Cornello M, Boscolo Brusà R, Sfriso A, Bonometto A. Changes in Macrozoobenthos Community after Aquatic Plant Restoration in the Northern Venice Lagoon (IT). International Journal of Environmental Research and Public Health. 2022; 19(8):4838. https://doi.org/10.3390/ijerph19084838
Chicago/Turabian StyleOselladore, Federica, Valentina Bernarello, Federica Cacciatore, Michele Cornello, Rossella Boscolo Brusà, Adriano Sfriso, and Andrea Bonometto. 2022. "Changes in Macrozoobenthos Community after Aquatic Plant Restoration in the Northern Venice Lagoon (IT)" International Journal of Environmental Research and Public Health 19, no. 8: 4838. https://doi.org/10.3390/ijerph19084838