
Interspecific Aggression and Food Competition between the Global Invader Palaemon macrodactylus and the Native Palaemon elegans


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Shrimp Collection and Housing
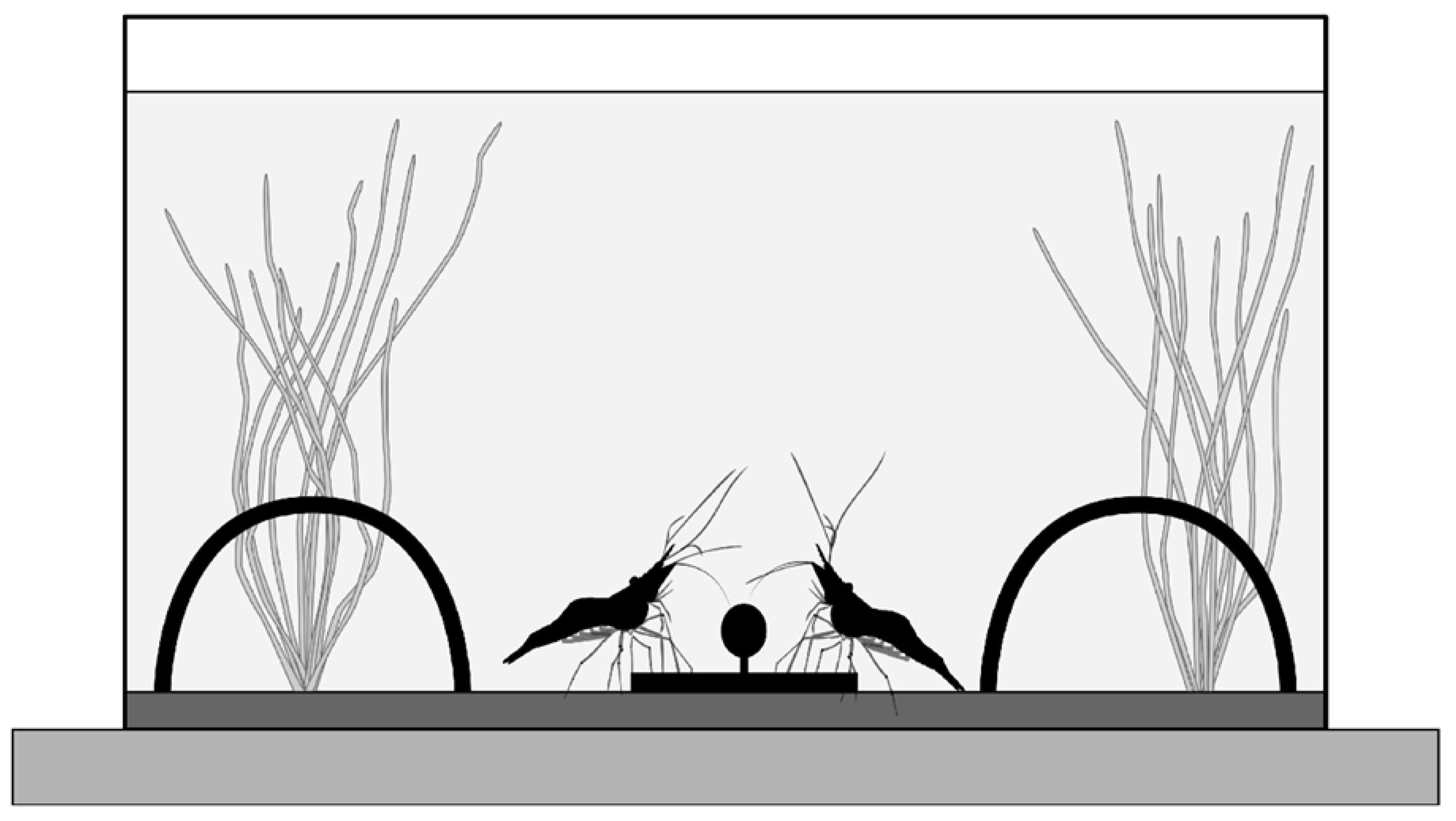
2.2. Experimental Apparatus and Procedure
2.3. Analysis of Behavioral Data
- -
- Latency time (LT): time in seconds (s) between the introduction of food and the first access to the resource. It is considered as access when the individual has begun moving towards the resource.
- -
- Residence time (RT): total time in seconds (s) spent for the consumption or protection of the food.
- -
- Number of chases (CN): the number of instantaneous aggressive behaviors enacted by an individual towards the other, as proposed by Eglė and Ulrika [22].
- -
- Number of escape (EN): number of tail-flip escapes after a chase or display.
2.4. Data Analysis
- -
- A t-test for dependent, paired observations was carried out to compare the inter-specific differences in frequency or duration of each behavioral parameter within each salinity condition.
- -
- GLM (Generalized Linear Model) was used with salinity and species as predictors together with the individuals’ weight as covariate.
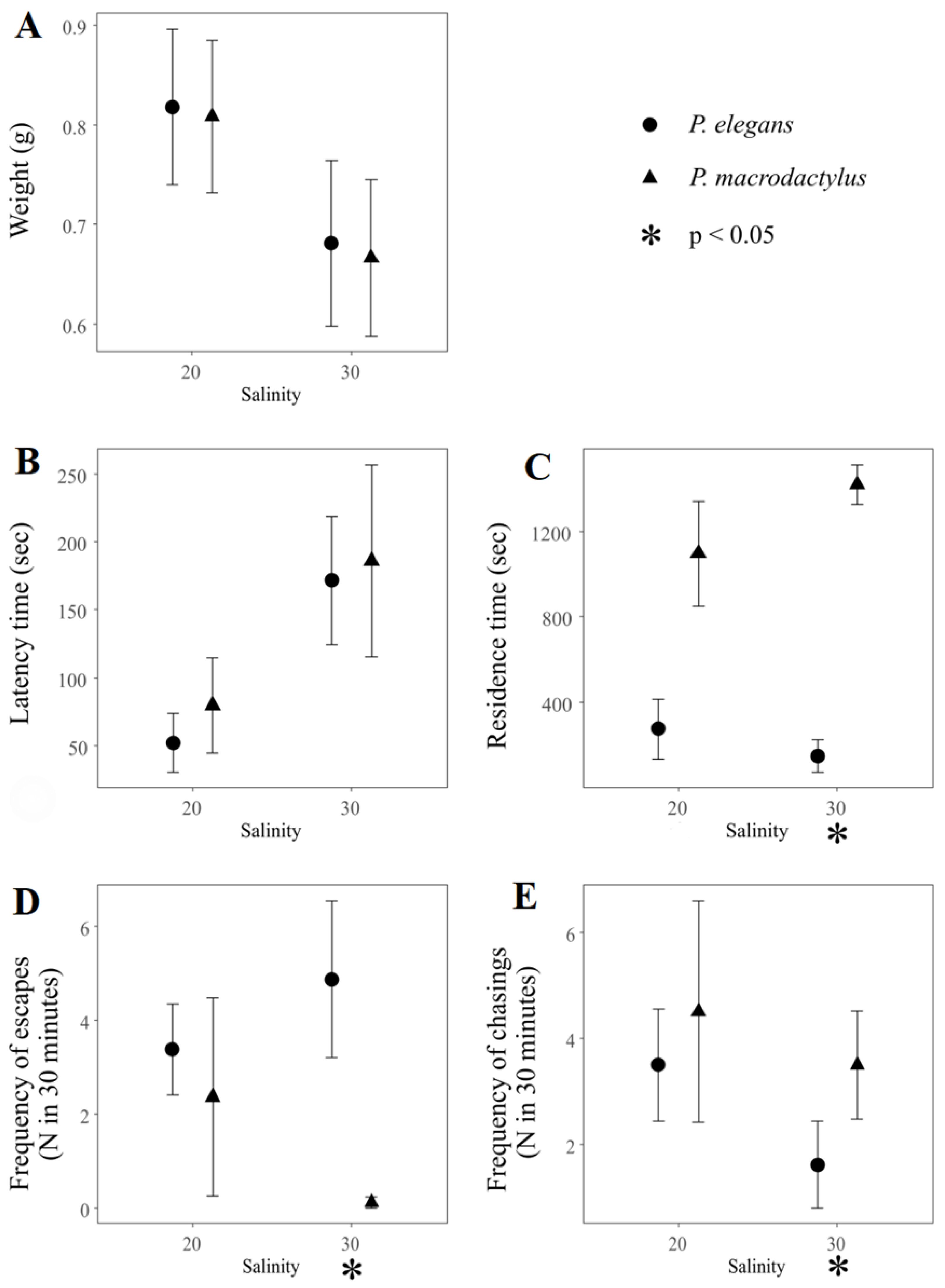
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean–the 100 “Worst Invasives” and their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef] [Green Version]
- Corriero, G.; Pierri, C.; Accoroni, S.; Alabiso, G.; Bavestrello, G.; Barbone, E.; Bastianini, M.; Bazzoni, A.M.; Bernardi Aubry, F.; Boero, F.; et al. Ecosystem vulnerability to alien and invasive species: A case study on marine habitats along the Italian coast. Aquat. Conserv.: Mar. Freshw. Ecosyst. 2016, 26, 392–409. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M.C.; Giaccone, G.; et al. Alien species along the Italian coasts: An overview. Biol. Invasions 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Rodríguez, G.; Suárez, H. Anthropogenic dispersal of decapod crustaceans in aquatic environments. Interciencia 2001, 26, 282–288. [Google Scholar]
- Ashelby, C.W.; De Grave, S.; Johnson, M.L. The global invader Palaemon macrodactylus (Decapoda, Palaemonidae): An interrogation of records and synthesis of data. Crustaceana 2013, 86, 594–624. [Google Scholar]
- AquaNIS Editorial Board. Information system on Aquatic Non-Indigenous and Cryptogenic Species. World Wide Web electronic publication. Version 2.36+. 2015. Available online: http://www.corpi.ku.lt/databases/aquanis (accessed on 4 October 2022).
- Lejeusne, C.; Saunier, A.; Petit, N.; Béguer, M.; Otani, M.; Carlton, J.T.; Rico, C.; Green, A.J. High genetic diversity and absence of founder effects in a worldwide aquatic invader. Sci. Rep. 2014, 4, 5808. [Google Scholar] [CrossRef] [Green Version]
- Cavraro, F.; Facca, C.; Naseer, M.; Malvasi, S. Comparing the reproductive success of three Palaemonid species in a Mediterranean coastal lagoon: Native and invasive responses to salinity changes. Hydrobiologia 2022, 849, 661–674. [Google Scholar] [CrossRef]
- Chícharo, M.A.; Leitão, T.; Range, P.; Gutierrez, C.; Morales, J.; Morais, P.; Chicharo, L. Alien species in the Guadiana Estuary (SE-Portugal/SW-Spain): Blackfordia virginica (Cnidaria, Hydrozoa) and Palaemon macrodactylus (Crustacea, Decapoda): Potential impacts and mitigation measures. Aquat. Invasions 2009, 4, 501–506. [Google Scholar] [CrossRef]
- González-Ortegón, E.; Pascual, E.; Cuesta, J.A.; Drake, P. Field distribution and osmoregulatory capacity of shrimps in a temperate European estuary (SW Spain). Estuar. Coast. Shelf S 2006, 67, 293–302. [Google Scholar] [CrossRef]
- González-Ortegón, E.; Cuesta, J.A.; Pascual, E.; Drake, P. Assessment of the interaction between the white shrimp, Palaemon longirostris, and the exotic oriental shrimp, Palaemon macrodactylus, in a European estuary (SW Spain). Biol. Invasions 2010, 12, 1731–1745. [Google Scholar] [CrossRef]
- Lejeusne, C.; Latchere, O.; Petit, N.; Rico, C.; Green, A.J. Do invaders always perform better? Comparing the response of native and invasive shrimps to temperature and salinity gradients in south-west Spain. Estuar. Coast. Shelf S 2014, 136, 102–111. [Google Scholar] [CrossRef]
- Vázquez, M.G.; Bas, C.C.; Spivak, E.D. Ontogeny of salinity tolerance in the invasive shrimp Palaemon macrodactylus (Caridea: Palaemonidae). J. Crustac. Biol. 2016, 36, 214–219. [Google Scholar] [CrossRef]
- Vázquez, M.G.; Bas, C.C. Population distribution and group structure of the invasive shrimp Palaemon macrodactylus (Caridea: Palaemonidae) living in a fouling community. Mar. Biol. Res. 2019, 15, 282–289. [Google Scholar] [CrossRef]
- Ricketts, E.F.; Calvin, J.; Hedgpeth, J.W. Between Pacific Tides; Stanford University Press: Redwood City, CA, USA, 1968. [Google Scholar]
- Ashelby, C.W.; De Grave, S.; Johnson, M.L. Diet analysis indicates seasonal fluctuation in trophic overlap and separation between a native abd an introduced shrimp species (Decapoda, Palaemonida) in the tidal river Thames (U.K.). Crustaceana 2016, 89, 701–719. [Google Scholar] [CrossRef]
- Evans, D.L.; Shehadi-Moacdieh, M. Body size and prior residency in staged encounters between female prawns, Palaemon elegans Rathke (Decapoda: Palaemonidae). Anim. Behav. 1988, 36, 452–455. [Google Scholar] [CrossRef]
- Nielsen, A.; Hagerman, L. Effects of short-term hypoxia on metabolism and haemocyanin oxygen transport in the prawns Palaemon adspersus and Palaemonetes varians. Mar. Ecol. Prog. Ser. 1998, 167, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Béguer, M.; Girardin, M.; Boet, P. First record of the invasive oriental shrimp Palaemon macrodactylus Rathbun, 1902 in France (Gironde Estuary). Aquat. Invasions 2007, 2, 132–136. [Google Scholar] [CrossRef]
- Correa, C.; Thiel, M. Mating systems in caridean shrimp (Decapoda: Caridea) and their evolutionary consequences for sexual dimorphism and reproductive biology. Rev. Chil. Hist. Nat. 2003, 76, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Cavraro, F.; Zucchetta, M.; Franzoi, P. First record of adult individuals of the Oriental shrimp Palaemon macrodactylus Rathbun, 1902 in the Venice Lagoon (north Adriatic Sea, Italy). Bioinvasions Rec. 2014, 3, 269–273. [Google Scholar] [CrossRef]
- Jakubavičiūtė, E.; Candolin, U. Density-dependent behavioural interactions influence coexistence between a native and a non-native mesopredator. Biol. Invasions 2021, 23, 3427–3434. [Google Scholar] [CrossRef]
- Chavanich, S.; Viyakarn, V.; Senanan, W.; Panutrakul, S. Laboratory assessment of feeding-behavior interactions between the introduced Pacific white shrimp Litopenaeus vannamei (Boone, 1931) (Penaeidae) and five native shrimps plus a crab species in Thailand. Aquat. Invasions 2016, 11, 67–74. [Google Scholar] [CrossRef]
- Vázquez, M.G.; Ituarte, R.B.; Bas, C.C.; Spivak, E.D. Effects of temperature and salinity on the ovarian cycle and the embryonic development of the invasive shrimp Palaemon macrodactylus. J. Crustacean Biol. 2013, 33, 218–223. [Google Scholar] [CrossRef]
- Redolfi Bristol, S.; Scapin, L.; Cavraro, F.; Facca, C.; Zucchetta, M.; Franzoi, P. Distribution of the alien species Palaemon macrodactylus Rathbun, 1902 in the Venice lagoon. It. J. Fresh Ichthyol. 2021, 7, 14–26. [Google Scholar]
- Janas, U.; Pilka, M.; Lipińska, D. Temperature and salinity requirements of Palaemon adspersus Rathke,1837 and Palaemon elegans Rathke, 1837. Do they explain the occurrence and expansion of prawns in the Baltic Sea? Mar. Biol. Res. 2013, 9, 293–300. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavraro, F.; Facca, C.; Altavilla, L.; Malavasi, S. Interspecific Aggression and Food Competition between the Global Invader Palaemon macrodactylus and the Native Palaemon elegans. J. Mar. Sci. Eng. 2022, 10, 1593. https://doi.org/10.3390/jmse10111593
Cavraro F, Facca C, Altavilla L, Malavasi S. Interspecific Aggression and Food Competition between the Global Invader Palaemon macrodactylus and the Native Palaemon elegans. Journal of Marine Science and Engineering. 2022; 10(11):1593. https://doi.org/10.3390/jmse10111593
Chicago/Turabian StyleCavraro, Francesco, Chiara Facca, Luca Altavilla, and Stefano Malavasi. 2022. "Interspecific Aggression and Food Competition between the Global Invader Palaemon macrodactylus and the Native Palaemon elegans" Journal of Marine Science and Engineering 10, no. 11: 1593. https://doi.org/10.3390/jmse10111593