
Targeting Non-Coding RNAs for the Development of Novel Hepatocellular Carcinoma Therapeutic Approaches
 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Hepatocellular Carcinoma
Available Therapeutic Approaches for HCC
2. Development of Novel Targeted Therapies for HCC
3. Non-Coding RNAs
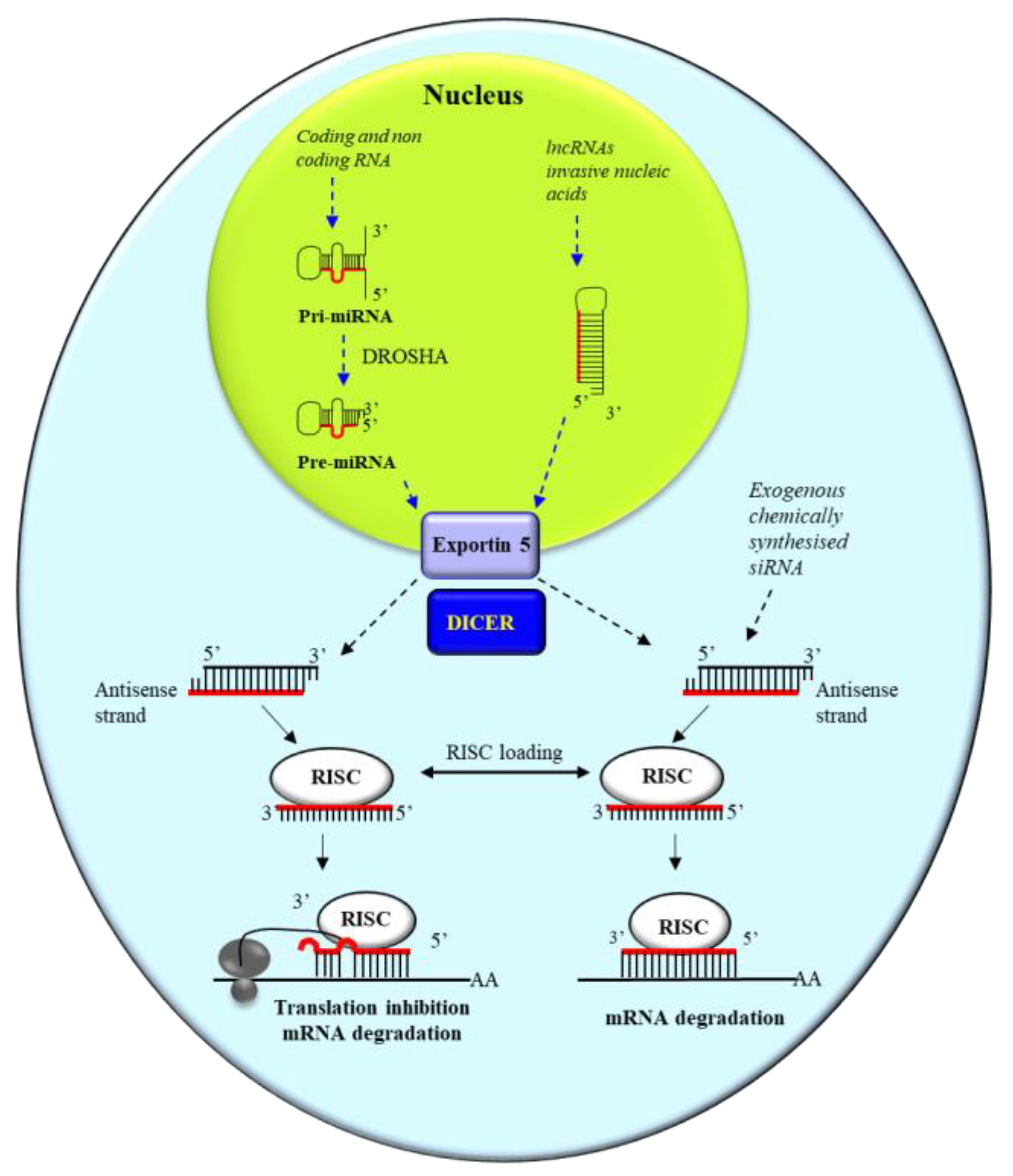
3.1. miRNA and siRNA
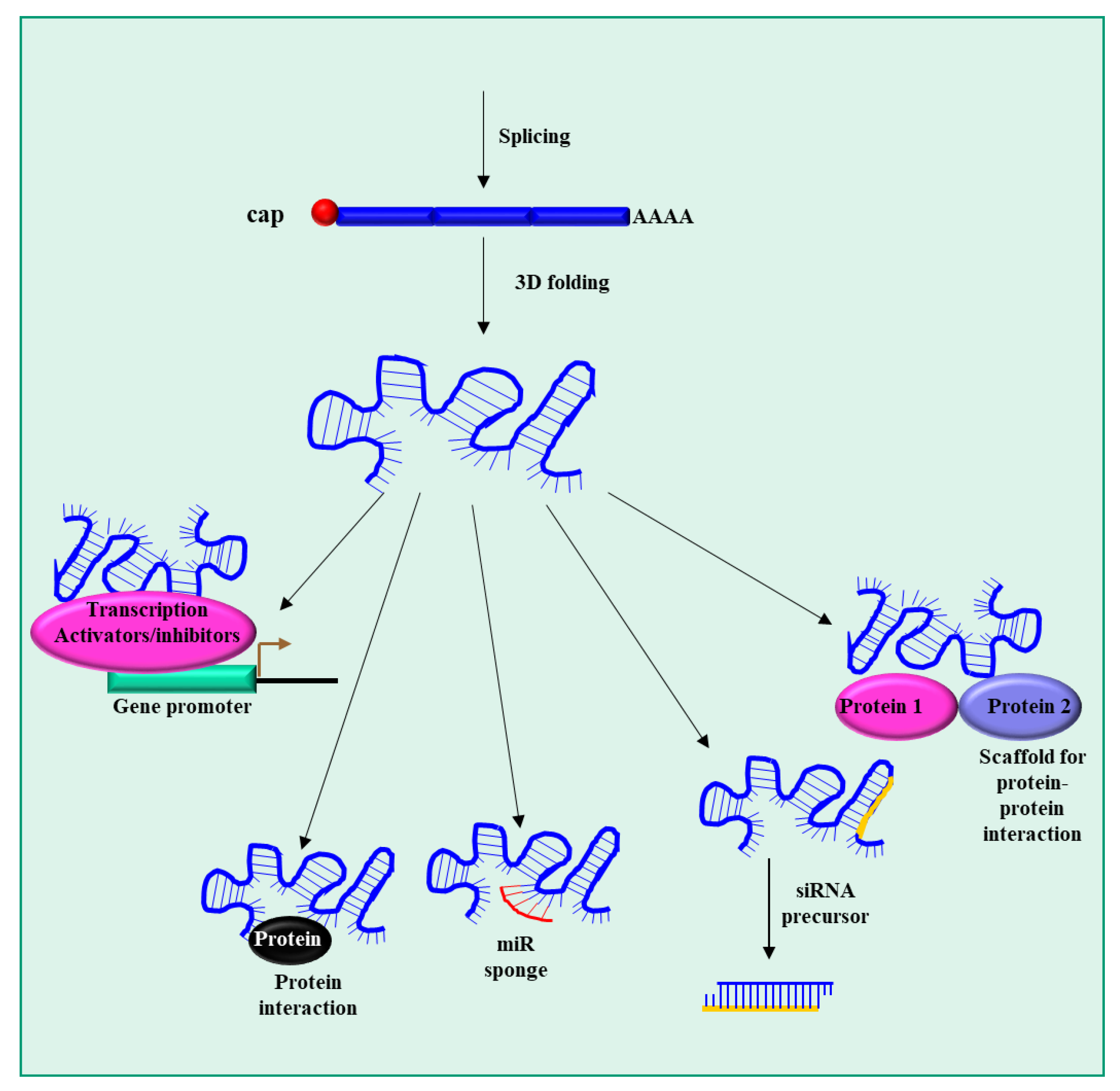
3.2. lncRNAs
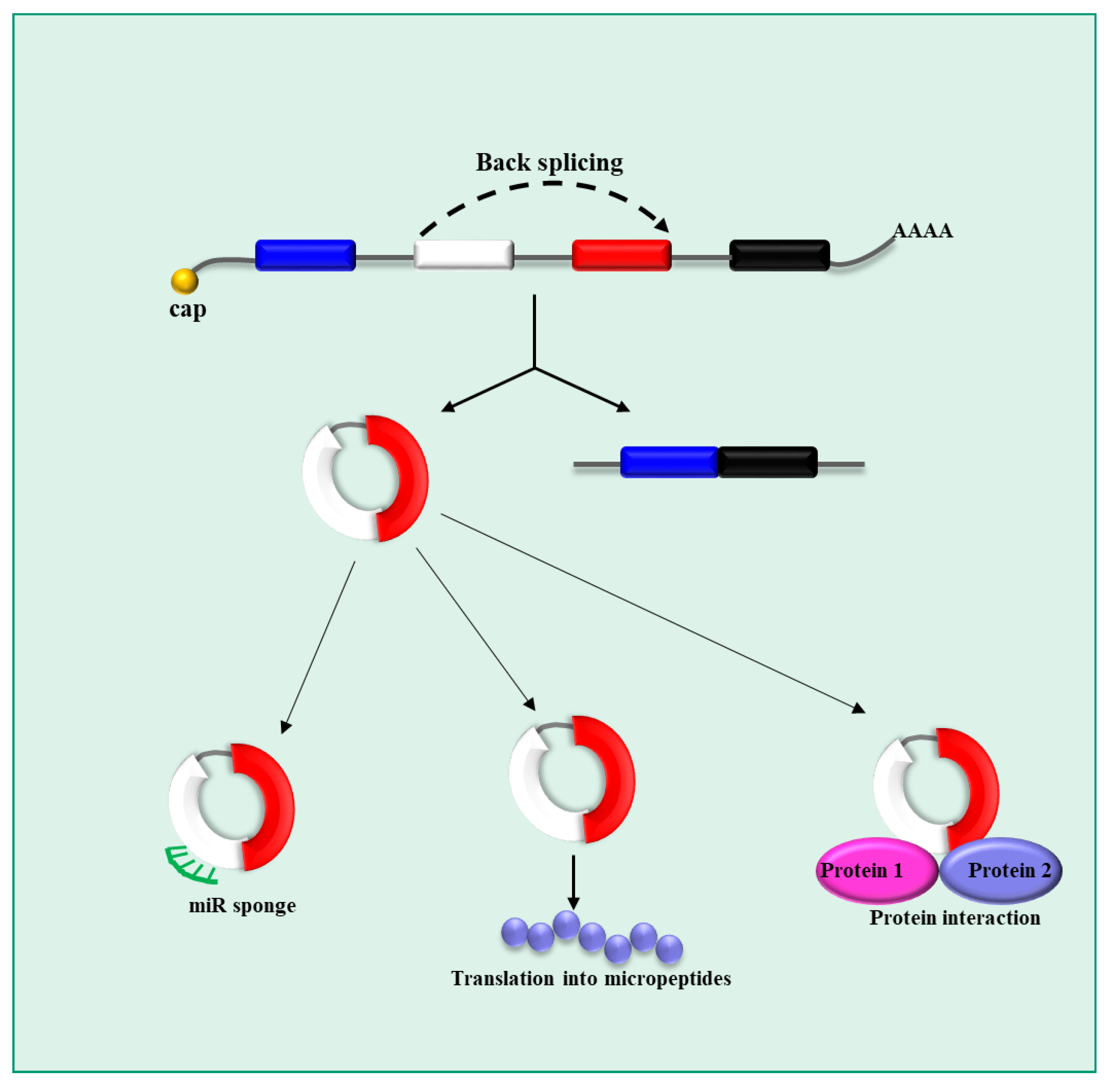
3.3. circRNAs
4. Therapeutic Potential of ncRNA Targeting
4.1. miRNAs
4.2. lncRNAs
4.3. circRNAs
4.4. ncRNAs and Drug Resistance
4.5. ncRNAs in Liver Fibrosis and HCC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dasgupta, P.; Henshaw, C.; Youlden, D.R.; Clark, P.J.; Aitken, J.F.; Baade, P.D. Global Trends in Incidence Rates of Primary Adult Liver Cancers: A Systematic Review and Meta-Analysis. Front Oncol. 2020, 10, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Petrick, J.L.; Florio, A.A.; Znaor, A.; Ruggieri, D.; Laversanne, M.; Alvarez, C.S.; Ferlay, J.; Valery, P.C.; Bray, F.; McGlynn, K.A. International trends in hepatocellular carcinoma incidence, 1978–2012. Int. J. Cancer 2020, 147, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev Dis. Prim. 2021, 7, 6. [Google Scholar] [CrossRef]
- Affo, S.; Yu, L.X.; Schwabe, R.F. The Role of Cancer-Associated Fibroblasts and Fibrosis in Liver Cancer. Annu. Rev Pathol. 2017, 12, 153–186. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Goto, T.; Hirotsu, Y.; Moriyama, M.; Omata, M. Molecular Mechanisms Driving Progression of Liver Cirrhosis towards Hepatocellular Carcinoma in Chronic Hepatitis B and C Infections: A Review. Int. J. Mol. Sci. 2019, 20, 1358. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef]
- Filliol, A.; Saito, Y.; Nair, A.; Dapito, D.H.; Yu, L.X.; Ravichandra, A.; Bhattacharjee, S.; Affo, S.; Fujiwara, N.; Su, H.; et al. Opposing roles of hepatic stellate cell subpopulations in hepatocarcinogenesis. Nature 2022, 610, 356–365. [Google Scholar] [CrossRef]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular Mechanisms of Liver Fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef]
- Chidambaranathan-Reghupaty, S.; Fisher, P.B.; Sarkar, D. Hepatocellular carcinoma (HCC): Epidemiology, etiology and molecular classification. Adv. Cancer Res. 2021, 149, 1–61. [Google Scholar]
- Vogel, A.; Meyer, T.; Sapisochin, G.; Salem, R.; Saborowski, A. Hepatocellular carcinoma. Lancet 2022, 400, 1345–1362. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Deng, Q.; Lin, S.; Wang, Y.; Xu, G. Comparison of microwave ablation and radiofrequency ablation for hepatocellular carcinoma: A systematic review and meta-analysis. Int. J. Hyperth. 2019, 36, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trotovsek, B.; Hadzialjevic, B.; Cemazar, M.; Sersa, G.; Djokic, M. Laparoscopic electrochemotherapy for the treatment of hepatocellular carcinoma: Technological advancement. Front. Oncol. 2022, 12, 996269. [Google Scholar] [CrossRef]
- Trotovsek, B.; Djokic, M.; Cemazar, M.; Sersa, G. New era of electrochemotherapy in treatment of liver tumors in conjunction with immunotherapies. World J. Gastroenterol. 2021, 27, 8216–8226. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Castet, F.; Heikenwalder, M.; Maini, M.K.; Mazzaferro, V.; Pinato, D.J.; Pikarsky, E.; Zhu, A.X.; Finn, R.S. Immunotherapies for hepatocellular carcinoma. Nat. Rev. Clin. Oncol. 2022, 19, 151–172. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Jiang, L.; Chen, Y. Recent advances in systemic therapy for hepatocellular carcinoma. Biomark. Res. 2022, 10, 3. [Google Scholar] [CrossRef]
- Farra, R.; Musiani, F.; Perrone, F.; Cemazar, M.; Kamensek, U.; Tonon, F.; Abrami, M.; Rucigaj, A.; Grassi, M.; Pozzato, G.; et al. Polymer-Mediated Delivery of siRNAs to Hepatocellular Carcinoma: Variables Affecting Specificity and Effectiveness. Molecules 2018, 23, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losurdo, P.; de Manzini, N.; Palmisano, S.; Grassi, M.; Parisi, S.; Rizzolio, F.; Tierno, D.; Biasin, A.; Grassi, C.; Truong, N.H.; et al. Potential Application of Small Interfering RNA in Gastro-Intestinal Tumors. Pharmaceuticals 2022, 15, 1295. [Google Scholar] [CrossRef]
- Zhou, F.; Teng, F.; Deng, P.; Meng, N.; Song, Z.; Feng, R. Recent Progress of Nano-drug Delivery System for Liver Cancer Treatment. Anticancer Agents Med. Chem. 2018, 17, 1884–1897. [Google Scholar] [CrossRef]
- Tang, R.; Li, R.; Li, H.; Ma, X.L.; Du, P.; Yu, X.Y.; Ren, L.; Wang, L.L.; Zheng, W.S. Design of Hepatic Targeted Drug Delivery Systems for Natural Products: Insights into Nomenclature Revision of Nonalcoholic Fatty Liver Disease. ACS Nano 2021, 15, 17016–17046. [Google Scholar] [CrossRef] [PubMed]
- Perrone, F.; Craparo, E.F.; Cemazar, M.; Kamensek, U.; Drago, S.E.; Dapas, B.; Scaggiante, B.; Zanconati, F.; Bonazza, D.; Grassi, M.; et al. Targeted delivery of siRNAs against hepatocellular carcinoma-related genes by a galactosylated polyaspartamide copolymer. J. Control. Release 2021, 330, 1132–1151. [Google Scholar] [CrossRef] [PubMed]
- Scarabel, L.; Perrone, F.; Garziera, M.; Farra, R.; Grassi, M.; Musiani, F.; Russo, S.C.; Salis, B.; De, S.L.; Toffoli, G.; et al. Strategies to optimize siRNA delivery to hepatocellular carcinoma cells. Expert Opin. Drug Deliv. 2017, 14, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Tsang, F.H.; Ng, I.O. Non-coding RNAs in hepatocellular carcinoma: Molecular functions and pathological implications. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Li, S.Y.; Fang, J.H.; Zhu, Y.; Yang, J.E. Functional long non-coding RNAs in hepatocellular carcinoma. Cancer Lett. 2021, 500, 281–291. [Google Scholar] [CrossRef]
- Chen, B.; Dragomir, M.P.; Yang, C.; Li, Q.; Horst, D.; Calin, G.A. Targeting non-coding RNAs to overcome cancer therapy resistance. Signal Transduct. Target Ther. 2022, 7, 121. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. MicroRNA maturation: Stepwise processing and subcellular localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [Green Version]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Bracht, J.; Hunter, S.; Eachus, R.; Weeks, P.; Pasquinelli, A.E. Trans-splicing and polyadenylation of let-7 microRNA primary transcripts. RNA 2004, 10, 1586–1594. [Google Scholar] [CrossRef] [Green Version]
- Gwizdek, C.; Bertrand, E.; Dargemont, C.; Lefebvre, J.C.; Blanchard, J.M.; Singer, R.H.; Doglio, A. Terminal minihelix, a novel RNA motif that directs polymerase III transcripts to the cell cytoplasm. Terminal minihelix and RNA export. J. Biol. Chem. 2001, 276, 25910–25918. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Peters, L.; Meister, G. Argonaute proteins: Mediators of RNA silencing. Mol. Cell 2007, 26, 611–623. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahpour, A.; Mullen, A.C. Our emerging understanding of the roles of long non-coding RNAs in normal liver function, disease, and malignancy. JHEP. Rep. 2021, 3, 100177. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Huang, B.; Zhang, K.; Liu, M.; Xu, T. Long non-coding RNA in cervical cancer: From biology to therapeutic opportunity. Biomed. Pharmacother. 2020, 127, 110209. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L. The biogenesis and emerging roles of circular RNAs. Nat. Rev. Mol. Cell Biol. 2016, 17, 205–211. [Google Scholar] [CrossRef]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glazar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-type specific features of circular RNA expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Huge, N.; Reinkens, T.; Buurman, R.; Sandbothe, M.; Bergmann, A.; Wallaschek, H.; Vajen, B.; Stalke, A.; Decker, M.; Eilers, M.; et al. MiR-129-5p exerts Wnt signaling-dependent tumor-suppressive functions in hepatocellular carcinoma by directly targeting hepatoma-derived growth factor HDGF. Cancer Cell Int. 2022, 22, 192. [Google Scholar] [CrossRef] [PubMed]
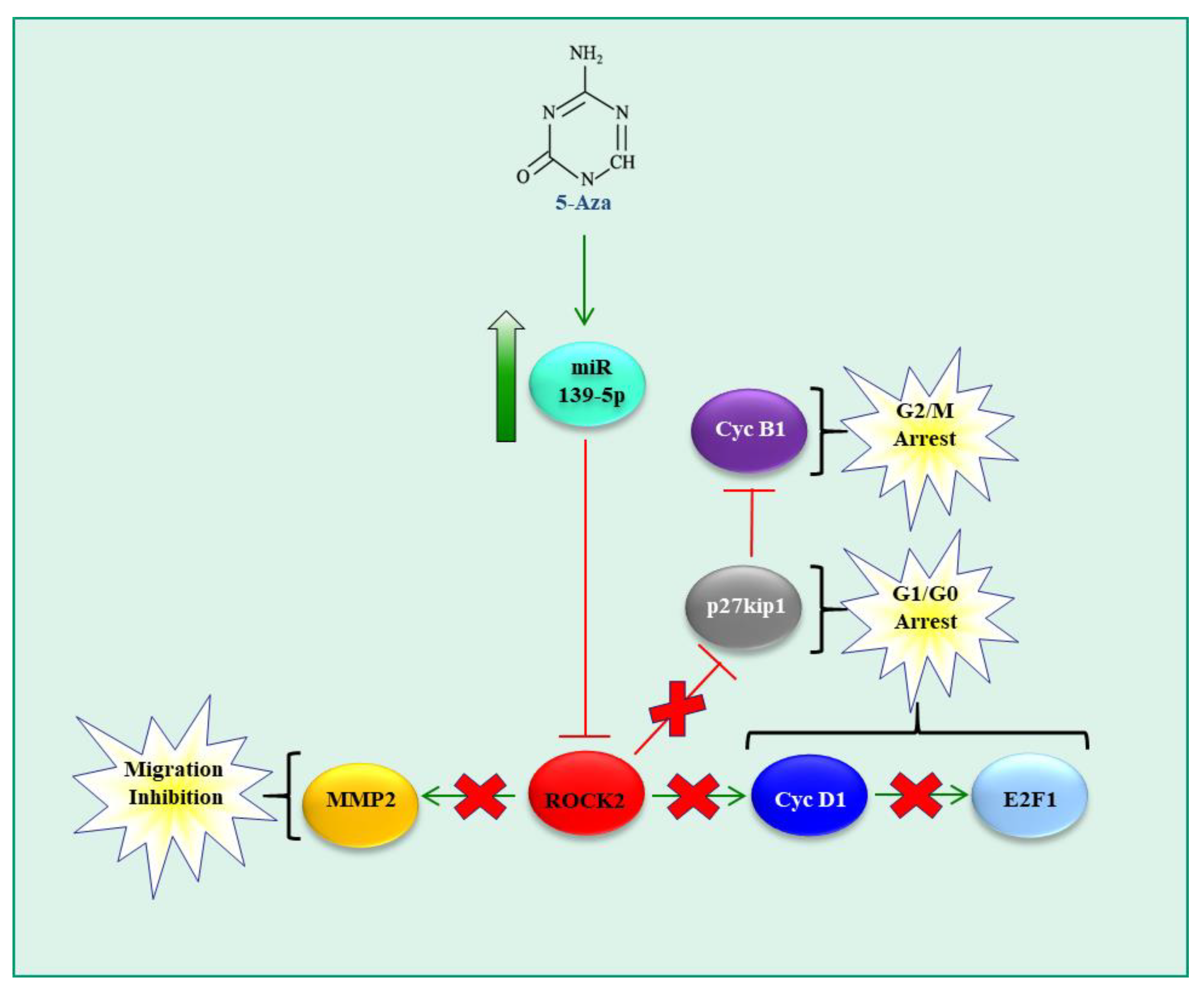
- Tonon, F.; Cemazar, M.; Kamensek, U.; Zennaro, C.; Pozzato, G.; Caserta, S.; Ascione, F.; Grassi, M.; Guido, S.; Ferrari, C.; et al. 5-Azacytidine Downregulates the Proliferation and Migration of Hepatocellular Carcinoma Cells In Vitro and In Vivo by Targeting miR-139-5p/ROCK2 Pathway. Cancers 2022, 14, 1630. [Google Scholar] [CrossRef]
- Gao, Y.; Gu, J.; Wang, Y.; Fu, D.; Zhang, W.; Zheng, G.; Wang, X. Hepatitis B virus X protein boosts hepatocellular carcinoma progression by downregulating microRNA-137. Pathol. Res. Pract. 2020, 216, 152981. [Google Scholar] [CrossRef]
- Zeng, C.; Yuan, G.; Hu, Y.; Wang, D.; Shi, X.; Zhu, D.; Hu, A.; Meng, Y.; Lu, J. Repressing phosphatidylinositol-4, 5-bisphosphate 3-kinase catalytic subunit gamma by microRNA-142-3p restrains the progression of hepatocellular carcinoma. Bioengineered 2022, 13, 1491–1506. [Google Scholar] [CrossRef]
- Kannan, M.; Jayamohan, S.; Moorthy, R.K.; Chabattula, S.C.; Ganeshan, M.; Arockiam, A.J.V. Dysregulation of miRISC Regulatory Network Promotes Hepatocellular Carcinoma by Targeting PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 11300. [Google Scholar] [CrossRef]
- Liang, Z.; Li, X.; Duan, F.; Song, L.; Wang, Z.; Li, X.; Yang, P.; Li, L. Protein tyrosine phosphatase non-receptor type 12 (PTPN12), negatively regulated by miR-106a-5p, suppresses the progression of hepatocellular carcinoma. Hum. Cell 2022, 35, 299–309. [Google Scholar] [CrossRef]
- Liu, Y.L.; Yang, W.H.; Chen, B.Y.; Nie, J.; Su, Z.R.; Zheng, J.N.; Gong, S.T.; Chen, J.N.; Jiang, D.; Li, Y. miR-29b suppresses proliferation and induces apoptosis of hepatocellular carcinoma ascites H22 cells via regulating TGF-β1 and p53 signaling pathway. Int. J. Mol. Med. 2021, 48, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Guo, C.; Liu, T.; Li, W.; Chen, X. MicroRNA miR-495 regulates the development of Hepatocellular Carcinoma by targeting C1q/tumor necrosis factor-related protein-3 (CTRP3). Bioengineered 2021, 12, 6902–6912. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Yang, Y.; Lin, C.; Wu, J.; Zhang, X. MiR-23a-3p promoted G1/S cell cycle transition by targeting protocadherin17 in hepatocellular carcinoma. J. Physiol. Biochem. 2020, 76, 123–134. [Google Scholar] [CrossRef]
- Zhao, Y.; Ye, L.; Yu, Y. MicroRNA-126-5p suppresses cell proliferation, invasion and migration by targeting EGFR in liver cancer. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Du, Y.; Li, H.; Huang, J.; Jiang, D.; Fan, J.; Shen, Y.; Zhang, L.; Yu, X.; Xu, N.; et al. miR-642 serves as a tumor suppressor in hepatocellular carcinoma by regulating SEMA4C and p38 MAPK signaling pathway. Oncol. Lett. 2020, 20, 74. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Zhang, T.; Hao, M.; Zhang, X.; Zhang, J.; Xie, Q.; Wang, Y.; Guo, M.; Zhuang, H.; et al. Methylation-mediated repression of microRNA 129-2 enhances oncogenic SOX4 expression in HCC. Liver Int. 2013, 33, 476–486. [Google Scholar] [CrossRef]
- Newell-Price, J.; Clark, A.J.; King, P. DNA methylation and silencing of gene expression. Trends Endocrinol. Metab. 2000, 11, 142–148. [Google Scholar] [CrossRef]
- Hendrich, B.; Bird, A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol. Cell Biol. 1998, 18, 6538–6547. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, K.; Steptoe, A.L.; Martin, H.C.; Pattabiraman, D.R.; Nones, K.; Waddell, N.; Mariasegaram, M.; Simpson, P.T.; Lakhani, S.R.; Vlassov, A.; et al. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA 2013, 19, 1767–1780. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gao, J.; Zhou, B.; Xie, J.; Zhou, G.; Chen, Y. Identification of prognostic markers for hepatocellular carcinoma based on miRNA expression profiles. Life Sci. 2019, 232, 116596. [Google Scholar] [CrossRef]
- Lu, A.Q.; Lv, B.; Qiu, F.; Wang, X.Y.; Cao, X.H. Upregulation of miR-137 reverses sorafenib resistance and cancer-initiating cell phenotypes by degrading ANT2 in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 2071–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, P.; Zhai, Z.; Hao, Z.; Zhang, H.; He, J. CircWHSC1 serves as an oncogene to promote hepatocellular carcinoma progression. Eur. J. Clin. Investig. 2021, 51, e13487. [Google Scholar] [CrossRef]
- Cui, D.; Ni, C. LncRNA Lnc712 Promotes Tumorigenesis in Hepatocellular Carcinoma by Targeting miR-142-3p/Bach-1 Axis. Cancer Manag. Res. 2020, 12, 11285–11294. [Google Scholar] [CrossRef]
- Ye, F.; Chen, C.; Qin, J.; Liu, J.; Zheng, C. Genetic profiling reveals an alarming rate of cross-contamination among human cell lines used in China. FASEB J. 2015, 29, 4268–4272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Zhuo, H.; Zhang, X.; Jiang, R.; Ji, J.; Deng, L.; Qian, X.; Zhang, F.; Sun, B. A novel biomarker Linc00974 interacting with KRT19 promotes proliferation and metastasis in hepatocellular carcinoma. Cell Death. Dis. 2014, 5, e1549. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hu, C.; Qu, X.; Chen, H.; Liu, L.; Zhou, L.; Liu, S.; Li, G.; Zhou, Y. Novel Role of Long Non-Coding RNA ASAP1-IT1 in Progression of Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 746896. [Google Scholar] [CrossRef]
- Pellegrino, R.; Castoldi, M.; Ticconi, F.; Skawran, B.; Budczies, J.; Rose, F.; Schwab, C.; Breuhahn, K.; Neumann, U.P.; Gaisa, N.T.; et al. LINC00152 Drives a Competing Endogenous RNA Network in Human Hepatocellular Carcinoma. Cells 2022, 11, 1528. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Fu, J.; Liu, Y.; Qu, W.; Wang, G.; Yan, Z. LncRNA CASC9 promotes proliferation, migration and inhibits apoptosis of hepatocellular carcinoma cells by down-regulating miR-424-5p. Ann. Hepatol. 2021, 23, 100297. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chang, Y.; Zhu, L.; Shen, C.; Qian, J.; Chang, R. LINC00839/miR-144-3p/WTAP (WT1 Associated protein) axis is involved in regulating hepatocellular carcinoma progression. Bioengineered 2021, 12, 10849–10861. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Yuan, J.; Zhang, N.; Zheng, Y.; Liu, J.; Yang, M. Silencing of lncRNA PVT1 by miR-214 inhibits the oncogenic GDF15 signaling and suppresses hepatocarcinogenesis. Biochem. Biophys. Res. Commun. 2020, 521, 478–484. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Sun, Q.C.; Zou, X.J.; Song, Y.; Li, W.W.; Guo, Z.Q.; Liu, S.S.; Liu, L.; Wu, D.H. Long noncoding RNA UPK1A-AS1 indicates poor prognosis of hepatocellular carcinoma and promotes cell proliferation through interaction with EZH2. J. Exp. Clin. Cancer Res. 2020, 39, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lou, G.; Li, A.; Liu, Y. lncRNA MALAT1 modulates cancer stem cell properties of liver cancer cells by regulating YAP1 expression via miR-375 sponging. Mol. Med. Rep. 2020, 22, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Nie, P.; Xu, S. Long noncoding RNA linc00467 plays an oncogenic role in hepatocellular carcinoma by regulating the miR-18a-5p/NEDD9 axis. J. Cell Biochem. 2020, 121, 3135–3144. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Chen, Z.; Gao, W.; Zhang, Y.; Wang, J.; Wang, J.; Cao, M.; Cai, J.; Wu, J.; Wang, X. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J. Hematol. Oncol. 2020, 13, 5. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Li, Q.; Zhi, L.J.; Li, J.M.; Wang, Z.Y. LINC00205 modulates the expression of EPHX1 through the inhibition of miR-184 in hepatocellular carcinoma as a ceRNA. J. Cell Physiol. 2020, 235, 3013–3021. [Google Scholar] [CrossRef]
- Tian, Q.; Yan, X.; Yang, L.; Liu, Z.; Yuan, Z.; Zhang, Y. Long non-coding RNA BACE1-AS plays an oncogenic role in hepatocellular carcinoma cells through miR-214-3p/APLN axis. Acta Biochim. Biophys. Sin. 2021, 53, 1538–1546. [Google Scholar] [CrossRef]
- Fu, Y.; Biglia, N.; Wang, Z.; Shen, Y.; Risch, H.A.; Lu, L.; Canuto, E.M.; Jia, W.; Katsaros, D.; Yu, H. Long non-coding RNAs, ASAP1-IT1, FAM215A, and LINC00472, in epithelial ovarian cancer. Gynecol. Oncol. 2016, 143, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Neumann, O.; Kesselmeier, M.; Geffers, R.; Pellegrino, R.; Radlwimmer, B.; Hoffmann, K.; Ehemann, V.; Schemmer, P.; Schirmacher, P.; Lorenzo, B.J.; et al. Methylome analysis and integrative profiling of human HCCs identify novel protumorigenic factors. Hepatology 2012, 56, 1817–1827. [Google Scholar] [CrossRef]
- Sharma, U.; Barwal, T.S.; Acharya, V.; Tamang, S.; Vasquez, K.M.; Jain, A. Cancer Susceptibility Candidate 9 (CASC9): A Novel Targetable Long Noncoding RNA in Cancer Treatment. Transl. Oncol. 2020, 13, 100774. [Google Scholar] [CrossRef]
- Yang, H.; Zheng, W.; Shuai, X.; Chang, R.M.; Yu, L.; Fang, F.; Yang, L.Y. MicroRNA-424 inhibits Akt3/E2F3 axis and tumor growth in hepatocellular carcinoma. Oncotarget 2015, 6, 27736–27750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Shen, H.; Zhu, X.; Liu, Y.; Yang, H.; Chen, H.; Xiong, S.; Chi, H.; Xu, W. A nuclear lncRNA Linc00839 as a Myc target to promote breast cancer chemoresistance via PI3K/AKT signaling pathway. Cancer Sci. 2020, 111, 3279–3291. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, C.; Chen, J.; Chen, D.; Yang, B.; He, B.; Hu, W.; Zhang, Y.; Liu, H.; Dai, L.; et al. WTAP facilitates progression of hepatocellular carcinoma via m6A-HuR-dependent epigenetic silencing of ETS1. Mol. Cancer 2019, 18, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Yuan, J.H.; Wang, S.B.; Yang, F.; Yuan, S.X.; Ye, C.; Yang, N.; Zhou, W.P.; Li, W.L.; Li, W.; et al. Oncofetal long noncoding RNA PVT1 promotes proliferation and stem cell-like property of hepatocellular carcinoma cells by stabilizing NOP2. Hepatology 2014, 60, 1278–1290. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, H.X.; Li, J.P.; Wang, S.; Fu, Z.; Jia, J.; Wang, L.; Zhu, Z.F.; Lu, R.; Yao, Z. Growth differentiation factor 15 induces growth and metastasis of human liver cancer stem-like cells via AKT/GSK-3beta/beta-catenin signaling. Oncotarget 2017, 8, 16972–16987. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Guo, T.; Cao, C. Restoration of UPK1A-AS1 Expression Suppresses Cell Proliferation, Migration, and Invasion in Esophageal Squamous Cell Carcinoma Cells Partially by Sponging microRNA-1248. Cancer Manag. Res. 2020, 12, 2653–2662. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y.; Mai, X.; Lu, S.; Jin, L.; Tai, X. STAT1-induced upregulation of LINC00467 promotes the proliferation migration of lung adenocarcinoma cells by epigenetically silencing DKK1 to activate Wnt/beta-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2019, 514, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Wang, Z.P.; Dang, Z.; Zheng, Z.G.; Li, X.; Zhou, L.; Ding, R.; Yue, S.Q.; Dou, K.F. Expression of NEDD9 in hepatocellular carcinoma and its clinical significance. Oncol. Rep. 2015, 33, 2375–2383. [Google Scholar] [CrossRef] [Green Version]
- Seitz, A.K.; Christensen, L.L.; Christensen, E.; Faarkrog, K.; Ostenfeld, M.S.; Hedegaard, J.; Nordentoft, I.; Nielsen, M.M.; Palmfeldt, J.; Thomson, M.; et al. Profiling of long non-coding RNAs identifies LINC00958 and LINC01296 as candidate oncogenes in bladder cancer. Sci. Rep. 2017, 7, 395. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.; Chen, Y.; Xie, Q.; Dong, N.; Gao, Y.; Deng, H.; Lu, C.; Wang, S. MicroRNA-214-3p inhibits proliferation and cell cycle progression by targeting MELK in hepatocellular carcinoma and correlates cancer prognosis. Cancer Cell Int. 2017, 17, 102. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wong, C.C.; Liu, D.; Go, M.Y.Y.; Wu, B.; Peng, S.; Kuang, M.; Wong, N.; Yu, J. APLN promotes hepatocellular carcinoma through activating PI3K/Akt pathway and is a druggable target. Theranostics 2019, 9, 5246–5260. [Google Scholar] [CrossRef] [PubMed]
- Du, A.; Li, S.; Zhou, Y.; Disoma, C.; Liao, Y.; Zhang, Y.; Chen, Z.; Yang, Q.; Liu, P.; Liu, S.; et al. M6A-mediated upregulation of circMDK promotes tumorigenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. Mol. Cancer 2022, 21, 109. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Zhang, B.Y.; Chen, Z.C. circRERE Regulates the Expression of GBX2 through miR-1299 and ZC3H13/N6-methyladenosine (m6A) to Promote Growth and Invasion of Hepatocellular Carcinoma Cells. J. Biosci. 2022, 47, 52. [Google Scholar] [CrossRef] [PubMed]
- Lei, D.; Wang, T. circSYPL1 Promotes the Proliferation and Metastasis of Hepatocellular Carcinoma via the Upregulation of EZH2 Expression by Competing with hsa-miR-506-3p. J. Oncol. 2022, 2022, 2659563. [Google Scholar] [CrossRef]
- Guo, X.; Wang, Z.; Deng, X.; Lu, Y.; Huang, X.; Lin, J.; Lan, X.; Su, Q.; Wang, C. Circular RNA CircITCH (has-circ-0001141) suppresses hepatocellular carcinoma (HCC) progression by sponging miR-184. Cell Cycle 2022, 21, 1557–1577. [Google Scholar] [CrossRef]
- Hu, H.; Wang, Y.; Qin, Z.; Sun, W.; Chen, Y.; Wang, J.; Wang, Y.; Nie, J.; Chen, L.; Cai, S.; et al. Regulation of MRP4 Expression by circHIPK3 via Sponging miR-124-3p/miR-4524-5p in Hepatocellular Carcinoma. Biomedicines 2021, 9, 497. [Google Scholar] [CrossRef]
- Wang, G.; Jiang, Y.; Lu, C.; Jiang, W.; Wu, S.; Hua, Y. CircFOXM1 promotes proliferation and metastasis of hepatocellular carcinoma via regulating miR-1179/SPAG5 axis. Sci Rep. 2021, 11, 23890. [Google Scholar] [CrossRef]
- Li, L.; Xiao, C.; He, K.; Xiang, G. Circ_0072088 promotes progression of hepatocellular carcinoma by activating JAK2/STAT3 signaling pathway via miR-375. IUBMB Life 2021, 73, 1153–1165. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, Y.; Jiang, H.Y.; Cheng, Z.M.; Wei, Z.J.; Wei, Y.H.; Liu, T.; Xia, B.J.; Zhao, X.Y.; Huang, Y.; et al. CircC16orf62 promotes hepatocellular carcinoma progression through the miR-138-5p/PTK2/AKT axis. Cell Death Dis. 2021, 12, 597. [Google Scholar] [CrossRef]
- Bao, S.; Jin, S.; Wang, C.; Tu, P.; Hu, K.; Lu, J. Androgen receptor suppresses vasculogenic mimicry in hepatocellular carcinoma via circRNA7/miRNA7-5p/VE-cadherin/Notch4 signalling. J. Cell Mol. Med. 2020, 24, 14110–14120. [Google Scholar] [CrossRef]
- Gao, J.; Li, E.; Liu, W.; Yang, Q.; Xie, C.; Ai, J.; Zhou, F.; Liao, W.; Wu, L. Circular RNA MYLK Promotes Hepatocellular Carcinoma Progression Through the miR29a/KMT5C Signaling Pathway. Onco. Targets Ther. 2020, 13, 8615–8627. [Google Scholar] [CrossRef]
- Pu, J.; Wang, J.; Li, W.; Lu, Y.; Wu, X.; Long, X.; Luo, C.; Wei, H. hsa_circ_0000092 promotes hepatocellular carcinoma progression through up-regulating HN1 expression by binding to microRNA-338-3p. J. Cell Mol. Med. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Sheng, P.; Sun, J.; Zhao, X.; Zhang, J.; Li, Y.; Zhang, Y.; Zhang, W.; Wang, J.; Liu, K.; et al. The circular RNA circMAST1 promotes hepatocellular carcinoma cell proliferation and migration by sponging miR-1299 and regulating CTNND1 expression. Cell Death Dis. 2020, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Song, J.; Tang, B.; Fang, S.; Zhang, D.; Zheng, L.; Wu, F.; Gao, Y.; Chen, C.; Hu, X.; et al. CircSOD2 induced epigenetic alteration drives hepatocellular carcinoma progression through activating JAK2/STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 259. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.H.; Wu, Q.W.; Li, X.D.; Wang, S.J.; Zhang, Z.M. SYPL1 overexpression predicts poor prognosis of hepatocellular carcinoma and associates with epithelial-mesenchymal transition. Oncol. Rep. 2017, 38, 1533–1542. [Google Scholar] [CrossRef] [Green Version]
- Lohr, N.J.; Molleston, J.P.; Strauss, K.A.; Torres-Martinez, W.; Sherman, E.A.; Squires, R.H.; Rider, N.L.; Chikwava, K.R.; Cummings, O.W.; Morton, D.H.; et al. Human ITCH E3 ubiquitin ligase deficiency causes syndromic multisystem autoimmune disease. Am. J. Hum. Genet. 2010, 86, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Wenzhi, G.; Jiakai, Z.; Dongyu, Z. Polymorphisms and Expression Pattern of Circular RNA Circ-ITCH Contributes to the Carcinogenesis of Hepatocellular Carcinoma. Oncotarget 2017, 28, 48169–48177. [Google Scholar]
- Porcelli, L.; Quatrale, A.E.; Mantuano, P.; Silvestris, N.; Brunetti, A.E.; Calvert, H.; Paradiso, A.; Azzariti, A. Synthetic lethality to overcome cancer drug resistance. Curr. Med. Chem. 2012, 19, 3858–3873. [Google Scholar] [CrossRef] [PubMed]
- Wierstra, I.; Alves, J. FOXM1, a typical proliferation-associated transcription factor. Biol. Chem. 2007, 388, 1257–1274. [Google Scholar] [CrossRef]
- Xiang, T.; Lin, Y.X.; Ma, W.; Zhang, H.J.; Chen, K.M.; He, G.P.; Zhang, X.; Xu, M.; Feng, Q.S.; Chen, M.Y.; et al. Vasculogenic mimicry formation in EBV-associated epithelial malignancies. Nat. Commun. 2018, 9, 5009. [Google Scholar] [CrossRef] [Green Version]
- Shao, B.; Zhao, X.; Liu, T.; Zhang, Y.; Sun, R.; Dong, X.; Liu, F.; Zhao, N.; Zhang, D.; Wu, L.; et al. LOXL2 promotes vasculogenic mimicry and tumour aggressiveness in hepatocellular carcinoma. J. Cell Mol. Med. 2019, 23, 1363–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Hu, Y.; Zeng, Q.; Wang, H.; Yan, J.; Li, H.; Yu, Z. Circular RNA MYLK promotes hepatocellular carcinoma progression by increasing Rab23 expression by sponging miR-362-3p. Cancer Cell Int. 2019, 19, 211. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Li, E.; Min, J.; Gong, C.; Gao, J.; Ai, J.; Liao, W.; Wu, L. miR-29a suppresses the growth and metastasis of hepatocellular carcinoma through IFITM3. Oncol. Rep. 2018, 40, 3261–3272. [Google Scholar] [CrossRef] [PubMed]
- Viotti, M.; Wilson, C.; McCleland, M.; Koeppen, H.; Haley, B.; Jhunjhunwala, S.; Klijn, C.; Modrusan, Z.; Arnott, D.; Classon, M.; et al. SUV420H2 is an epigenetic regulator of epithelial/mesenchymal states in pancreatic cancer. J. Cell Biol. 2018, 217, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Liu, G.; Huo, X.; Tao, X.; Sun, X.; Ge, Z.; Yang, J.; Fan, J.; Liu, L.; Qin, W. Hsa_circ_0001649: A circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark. 2016, 16, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Sun, X.; Mao, Q.Q.; Jiang, X.Y.; Zhao, X.G.; Xu, W.J.; Zhong, L. Increased expression of hematological and neurological expressed 1 (HN1) is associated with a poor prognosis of hepatocellular carcinoma and its knockdown inhibits cell growth and migration partly by down-regulation of c-Met. Kaohsiung. J. Med. Sci. 2020, 36, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, Z.; Zhang, Y.; Evert, M.; Calvisi, D.F.; Chen, X. beta-Catenin signaling in hepatocellular carcinoma. J. Clin. Investig. 2022, 132, e154515. [Google Scholar] [CrossRef]
- Luo, Z.; Mao, X.; Cui, W. Circular RNA expression and circPTPRM promotes proliferation and migration in hepatocellular carcinoma. Med. Oncol. 2019, 36, 86. [Google Scholar] [CrossRef]
- Lu, Y.; Chan, Y.T.; Tan, H.Y.; Zhang, C.; Guo, W.; Xu, Y.; Sharma, R.; Chen, Z.S.; Zheng, Y.C.; Wang, N.; et al. Epigenetic regulation of ferroptosis via ETS1/miR-23a-3p/ACSL4 axis mediates sorafenib resistance in human hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 3. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Chen, X.; Zhang, S.; Yun, Z.; Gao, Q.; Sheng, H.; Wang, J. Long non-coding RNA placenta-specific protein 2 regulates the chemosensitivity of cancer cells to cisplatin in hepatocellular carcinoma (HCC) by sponging microRNA-96 to upregulate X-linked inhibitor of apoptosis protein. Bioengineered 2022, 13, 10765–10773. [Google Scholar] [CrossRef]
- Chen, B.W.; Zhou, Y.; Wei, T.; Wen, L.; Zhang, Y.B.; Shen, S.C.; Zhang, J.; Ma, T.; Chen, W.; Ni, L.; et al. lncRNA-POIR promotes epithelial-mesenchymal transition and suppresses sorafenib sensitivity simultaneously in hepatocellular carcinoma by sponging miR-182-5p. J. Cell Biochem. 2021, 122, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Hirao, A.; Sato, Y.; Tanaka, H.; Nishida, K.; Tomonari, T.; Hirata, M.; Bando, M.; Kida, Y.; Tanaka, T.; Kawaguchi, T.; et al. MiR-125b-5p Is Involved in Sorafenib Resistance through Ataxin-1-Mediated Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma. Cancers 2021, 13, 4917. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, W.; Ye, Y.; Li, H.; Cheng, L.; Zhang, M.; Zheng, S.; Yu, J. LncRNA HOTAIR Contributes to Sorafenib Resistance through Suppressing miR-217 in Hepatic Carcinoma. Biomed. Res. Int. 2020, 2020, 9515071. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, Y.; Li, Z.; Li, H.; Li, X.; Yan, L.; Mao, J.; Shen, J.; Chen, W.; Xue, F. Long non-coding RNA H19 is involved in sorafenib resistance in hepatocellular carcinoma by upregulating miR-675. Oncol. Rep. 2020, 44, 165–173. [Google Scholar] [CrossRef]
- Yin, L.; Sun, T.; Liu, R. NACC-1 regulates hepatocellular carcinoma cell malignancy and is targeted by miR-760. Acta Biochim. Biophys. Sin. 2020, 52, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a new form of cell death: Opportunities and challenges in cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.J.; Cheng, D.; He, X.Y.; Meng, Z.; Li, W.Z.; Chen, R.F. Knockdown of Hotair suppresses proliferation and cell cycle progression in hepatocellular carcinoma cell by downregulating CCND1 expression. Mol. Med. Rep. 2017, 16, 4980–4986. [Google Scholar] [CrossRef] [Green Version]
- Gan, X.; Ding, D.; Wang, M.; Yang, Y.; Sun, D.; Li, W.; Ding, W.; Yang, F.; Zhou, W.; Yuan, S. DANCR deletion retards the initiation and progression of hepatocellular carcinoma based on gene knockout and patient-derived xenograft in situ hepatoma mice model. Cancer Lett. 2022, 550, 215930. [Google Scholar] [CrossRef]
- Yang, J.; Xu, C.; Wu, M.; Wu, Y.; Jia, X.; Zhou, C.; Zhang, X.; Ge, S.; Li, Z.; Zhang, L. MicroRNA-124 inhibits hepatic stellate cells inflammatory cytokines secretion by targeting IQGAP1 through NF-kappaB pathway. Int. Immunopharmacol. 2021, 95, 107520. [Google Scholar] [CrossRef]
- Feng, R.; Cui, Z.; Liu, Z.; Zhang, Y. Upregulated microRNA-132 in T helper 17 cells activates hepatic stellate cells to promote hepatocellular carcinoma cell migration in vitro. Scand. J. Immunol. 2021, 93, e13007. [Google Scholar] [CrossRef]
- Dong, Y.; Li, F.; Wang, J.; Hu, J.; Li, Z.; Gu, Y.; Feng, Y. miR-369 inhibits Liver Cancer progression by targeting ZEB1 pathway and predicts the prognosis of HCC patients. J. Cancer 2021, 12, 3067–3076. [Google Scholar] [CrossRef]
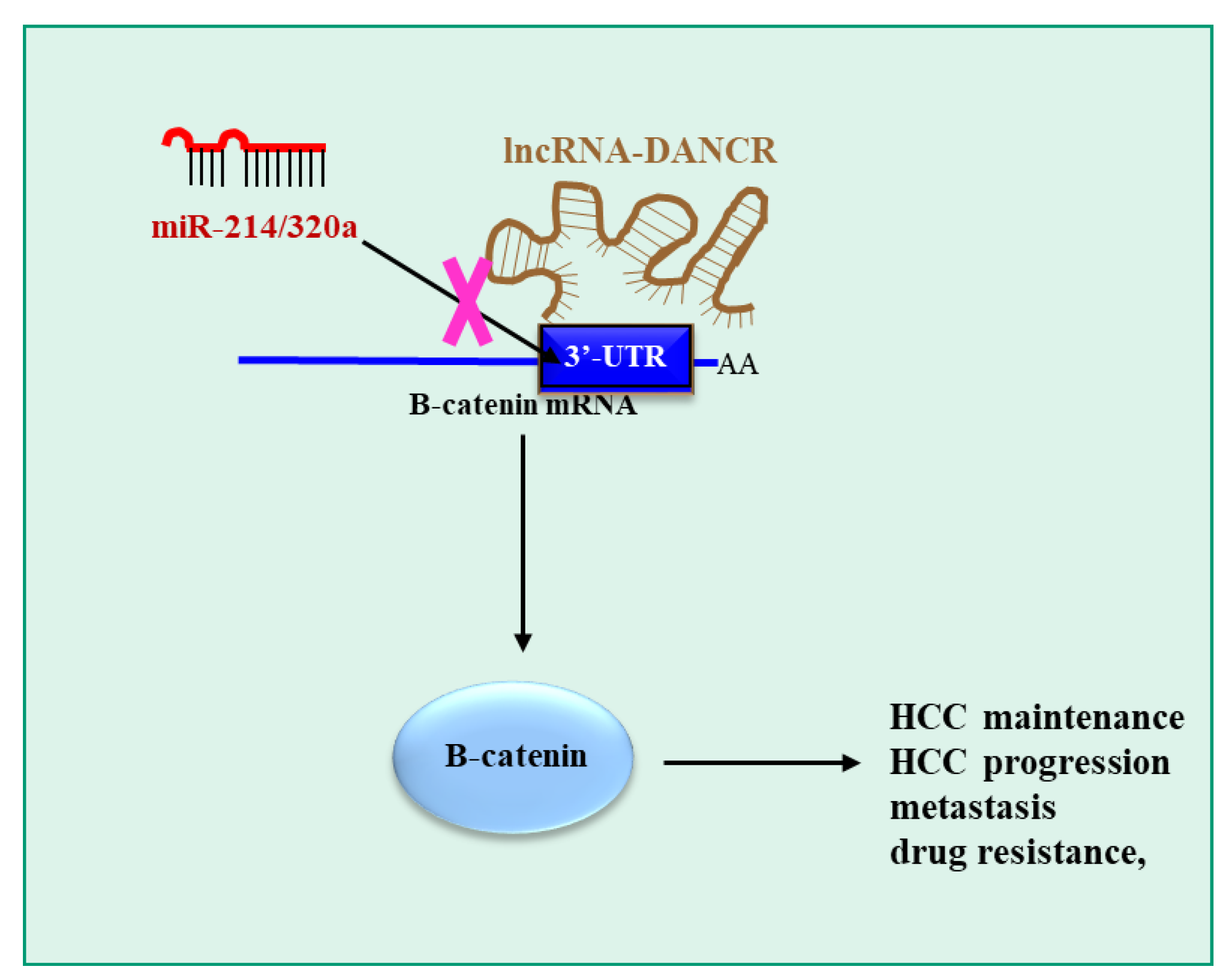
- Yuan, S.X.; Wang, J.; Yang, F.; Tao, Q.F.; Zhang, J.; Wang, L.L.; Yang, Y.; Liu, H.; Wang, Z.G.; Xu, Q.G.; et al. Long noncoding RNA DANCR increases stemness features of hepatocellular carcinoma by derepression of CTNNB1. Hepatology 2016, 63, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, V.; Turcios, L.; Marti, F.; Gedaly, R. Targeting Wnt/beta-catenin pathway in hepatocellular carcinoma treatment. World J. Gastroenterol. 2016, 22, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Long, H.D.; Ma, Y.S.; Yang, H.Q.; Xue, S.B.; Liu, J.B.; Yu, F.; Lv, Z.W.; Li, J.Y.; Xie, R.T.; Chang, Z.Y.; et al. Reduced hsa-miR-124-3p levels are associated with the poor survival of patients with hepatocellular carcinoma. Mol. Biol. Rep. 2018, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, L.; Liang, R.; Yang, C.; Wang, P. Baicalin suppresses renal fibrosis through microRNA-124/TLR4/NF-kappaB axis in streptozotocin-induced diabetic nephropathy mice and high glucose-treated human proximal tubule epithelial cells. J. Physiol. Biochem. 2020, 76, 407–416. [Google Scholar] [CrossRef]
- Zong, C.; Zhang, X.; Xie, Y.; Cheng, J. Transforming growth factor-beta inhibits IQ motif containing guanosine triphosphatase activating protein 1 expression in lung fibroblasts via the nuclear factor-kappaB signaling pathway. Mol. Med. Rep. 2015, 12, 442–448. [Google Scholar] [CrossRef]
- Qi, W.; Huang, X.; Wang, J. Correlation between Th17 cells and tumor microenvironment. Cell Immunol. 2013, 285, 18–22. [Google Scholar] [CrossRef]
- Grassi, G.; Marini, J.C. Ribozymes: Structure, function, and potential therapy for dominant genetic disorders. Ann. Med. 1996, 28, 499–510. [Google Scholar] [CrossRef]
- Grassi, G.; Schneider, A.; Engel, S.; Racchi, G.; Kandolf, R.; Kuhn, A. Hammerhead ribozymes targeted against cyclin E and E2F1 cooperate to down-regulate coronary smooth muscle cell proliferation. J. Gene Med. 2005, 7, 1223–1234. [Google Scholar] [CrossRef]
- Grassi, M.; Cavallaro, G.; Scirè, S.; Scaggiante, B.; Daps, B.; Farra, R.; Baiz, D.; Giansante, C.; Guarnieri, G.; Perin, D.; et al. Current Strategies to Improve the Efficacy and the Delivery of Nucleic Acid Based Drugs. Curr. Signal Transduct. Ther. 2010, 5, 92–120. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | miRNA Target * | Reference |
|---|---|---|
| miR-129-5p (Ts) | SOX4/HDGF | [46] |
| miR-139-5p (Ts) | ROCK2 | [47] |
| miR-137 (Ts) | Notch-1 | [48] |
| miR-142-3p (Ts) | PIK3CG | [49] |
| miR-221 (O) | Not indicated | [50] |
| miR-106a-5p (O) | PTPN12 | [51] |
| miR-29b-3p (Ts) | Not indicated | [52] |
| miR-495 (Ts) | CTRP3 | [53] |
| miR-23a-3p (O) | PCDH17 | [54] |
| miR-126-5p (Ts) | EGFR | [55] |
| miR-642a (O) | SEMA4C | [56] |
| lncRNA * | miRNA | miRNA Target * | Reference |
|---|---|---|---|
| lnc712 (O) | miR-142-3p-Ts | Bach-1 | [64] |
| lncRNA ASAP1-IT1 (O) | miR-221-3p-Ts | Not indicated | [67] |
| LINC00152 (O) | miR143a-3p-Ts | KLC2 | [68] |
| lncRNA CASC9 (O) | miR-424-5p-Ts | Not indicated | [69] |
| LINC00839 (O) | miR-144-3p-Ts | WTAP | [70] |
| lncPVT1 (O) | miR-214-Ts | GDF15 | [71] |
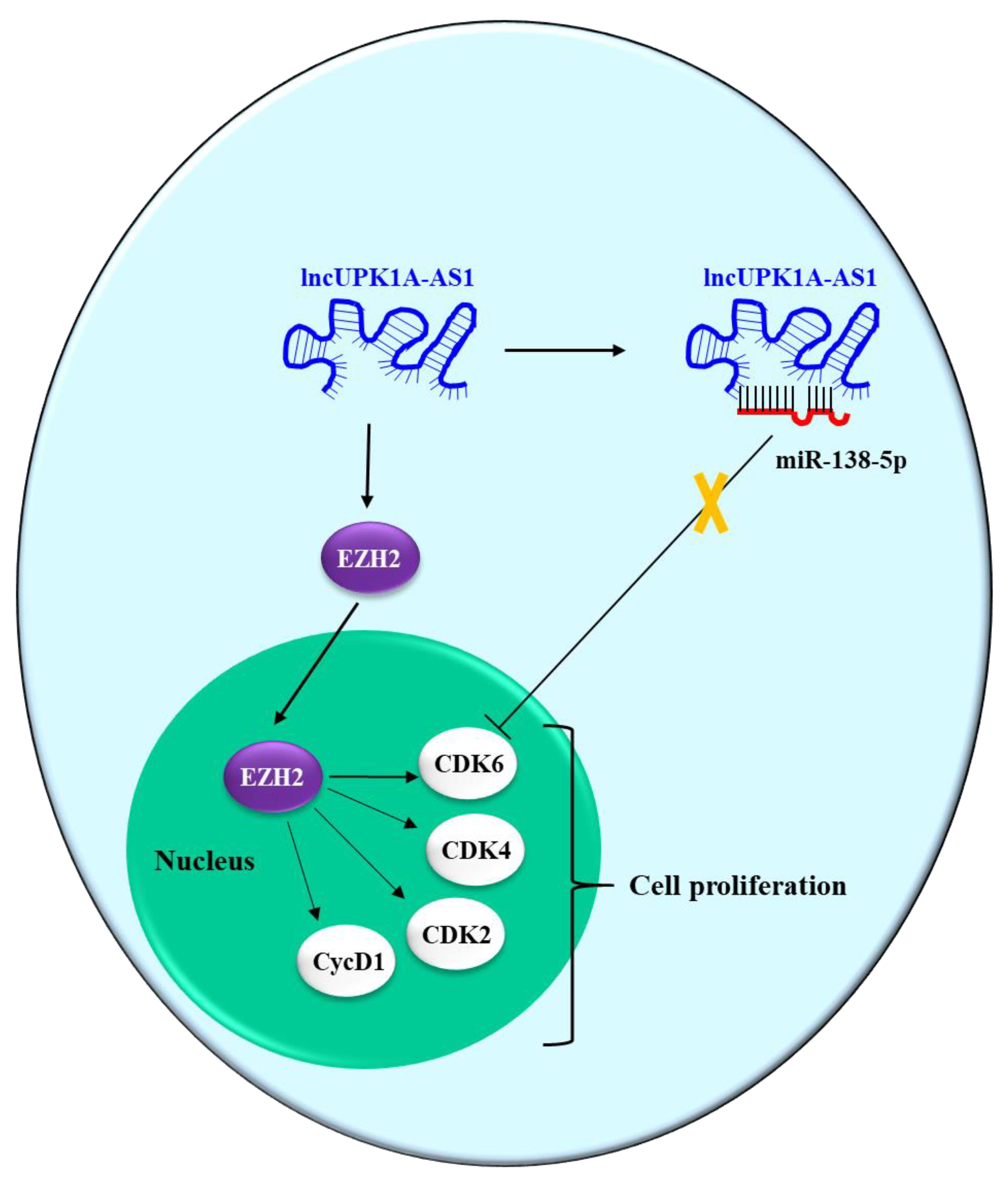
| lncUPK1A-AS1 (O) | miR-138-5p-Ts | CDK6 | [72] |
| lnc MALAT1 (O) | miR-375-Ts | YAP | [73] |
| lnc linc00467 (O) | miR-18a-5p-Ts | NEDD9 | [74] |
| lnc LINC00958-O (O) | miR-3619-5p-Ts | HDGF | [75] |
| lnc LINC00205 (Ts) | miR-184-O | EPHX1 | [76] |
| lnc BACE1-AS (O) | miR-214-3p-Ts | APLN | [77] |
| circRNA * | Sponged miRNA | miRNA Target * | Reference |
|---|---|---|---|
| circMDK (O) | miR-346, miR-874-3p-Ts | ATG19L1 | [92] |
| circRERE (O) | miR-1299-Ts | GBX2 | [93] |
| circSYPL1 (O) | miR-506-3p-Ts | EZH2 | [94] |
| circITCH (Ts) | miR-184-O | Not indicated | [95] |
| circHIPK3 (O) | miR-124-3p, miR-4524-5p-Ts | MRP4 | [96] |
| circFOXM1 (O) | miR-1179-Ts | SPAG5 | [97] |
| circ0072088 (O) | miR-375-Ts | JAK2 | [98] |
| circC16orf62 (O) | miR-138-5p-Ts | PTK2 | [99] |
| circRNA7 (O) | miR-7-5p-Ts | VEcadherin/Notch | [100] |
| circMYLK (O) | miR-29a-Ts | KMT5C | [101] |
| circ0000092 (O) | miR-338-3p-Ts | HN1 | [102] |
| circMAST1 (O) | miR-1299-Ts | CTNNB1 | [103] |
| circSOD2 (O) | miR-502-5p-Ts | DNMT3a | [104] |
| CircWHSC1 (O) | miR-142-3-Ts | HOXA1 | [63] |
| lncRNA * | miRNA | miRNA Target * | Reference |
|---|---|---|---|
| miR-23a-3p (O) | ACSL4 | [119] | |
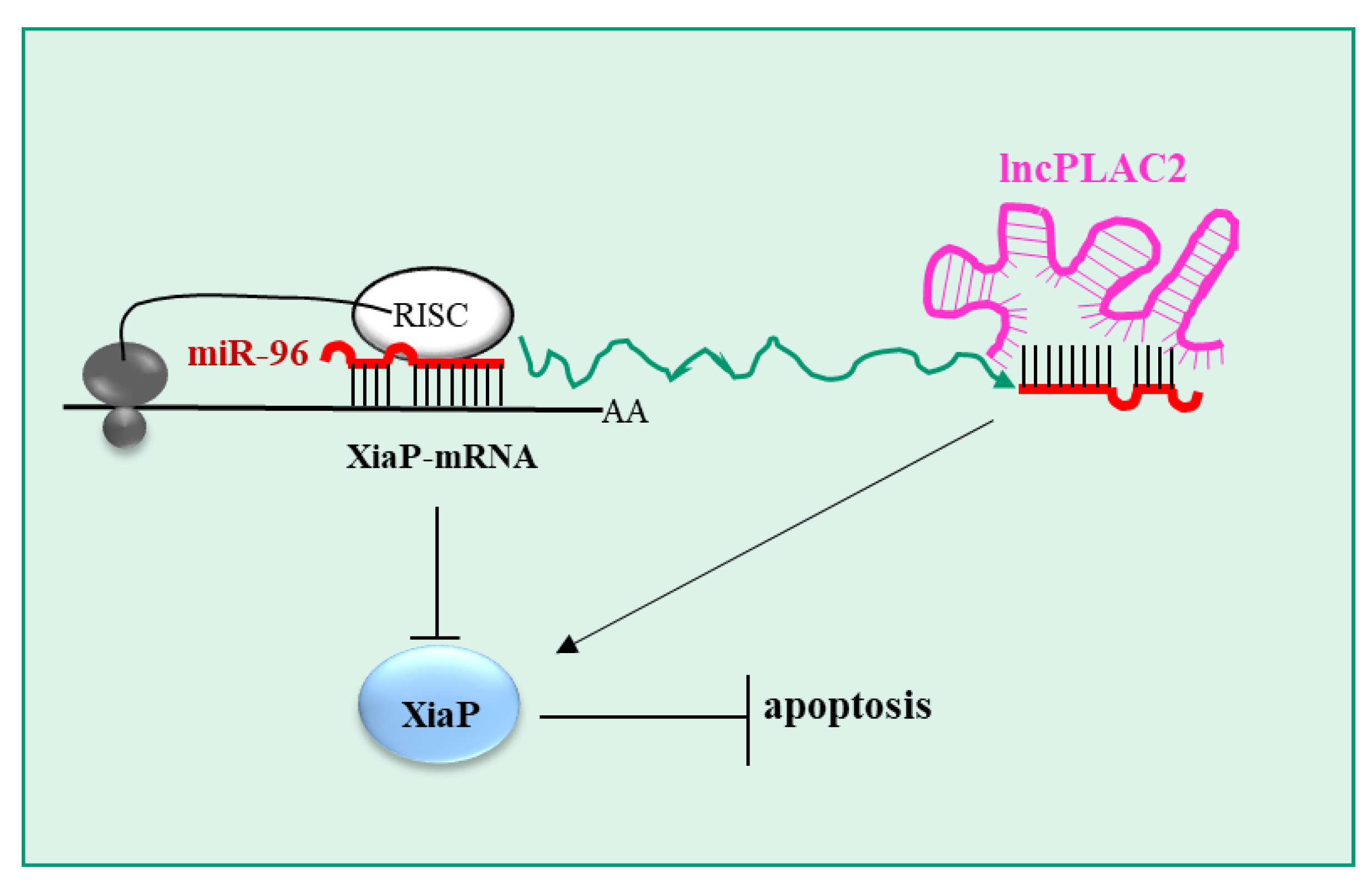
| lncRNA-PLAC2 (O) | miR-96 (Ts) | XiaP | [120] |
| lncRNA-POIR (O) | miR-182-5p (Ts) | Vimentin? | [121] |
| miR-125b-5p (O) | ATXN1 | [122] | |
| lncRNA-HOTAIR (O) | miR-217 (Ts) | Vimentin? | [123] |
| lncRNA-H19 (O) | miR-675 (O) | Not indicated | [124] |
| miR-760 (Ts) | NACC-1 | [125] |
| lncRNA * | miRNA | miRNA Target * | Reference |
|---|---|---|---|
| lncRNA-DANCR (O) | miR-214, miR-320a (Ts) | CTNNB1 | [128] |
| miR-124 (O) | IQGAP1 | [129] | |
| miR-132 | SNIP1 | [130] | |
| miR-369 (Ts) | ZEB1 | [131] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesenko, T.; Brezar, S.K.; Cemazar, M.; Biasin, A.; Tierno, D.; Scaggiante, B.; Grassi, M.; Grassi, C.; Dapas, B.; Truong, N.H.; et al. Targeting Non-Coding RNAs for the Development of Novel Hepatocellular Carcinoma Therapeutic Approaches. Pharmaceutics 2023, 15, 1249. https://doi.org/10.3390/pharmaceutics15041249
Jesenko T, Brezar SK, Cemazar M, Biasin A, Tierno D, Scaggiante B, Grassi M, Grassi C, Dapas B, Truong NH, et al. Targeting Non-Coding RNAs for the Development of Novel Hepatocellular Carcinoma Therapeutic Approaches. Pharmaceutics. 2023; 15(4):1249. https://doi.org/10.3390/pharmaceutics15041249
Chicago/Turabian StyleJesenko, Tanja, Simona Kranjc Brezar, Maja Cemazar, Alice Biasin, Domenico Tierno, Bruna Scaggiante, Mario Grassi, Chiara Grassi, Barbara Dapas, Nhung Hai Truong, and et al. 2023. "Targeting Non-Coding RNAs for the Development of Novel Hepatocellular Carcinoma Therapeutic Approaches" Pharmaceutics 15, no. 4: 1249. https://doi.org/10.3390/pharmaceutics15041249