
Molecular and Morphometric Update on Italian Salicornia (Chenopodiaceae), with a Focus on the Species S. procumbens s. l.

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
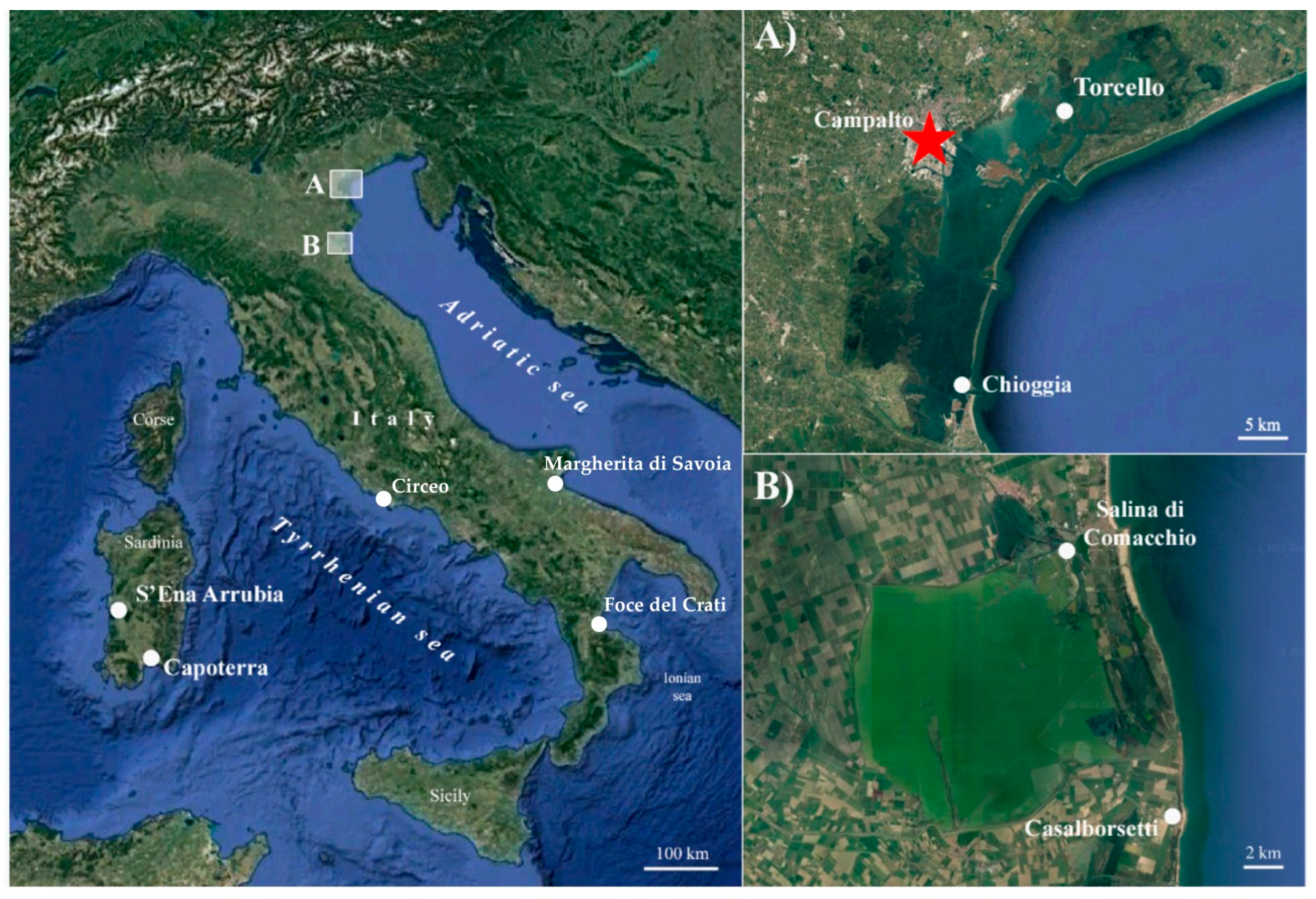
2.1. Plant Material
2.2. Molecular and Phylogenetic Analyses
2.3. Morphometric Analyses
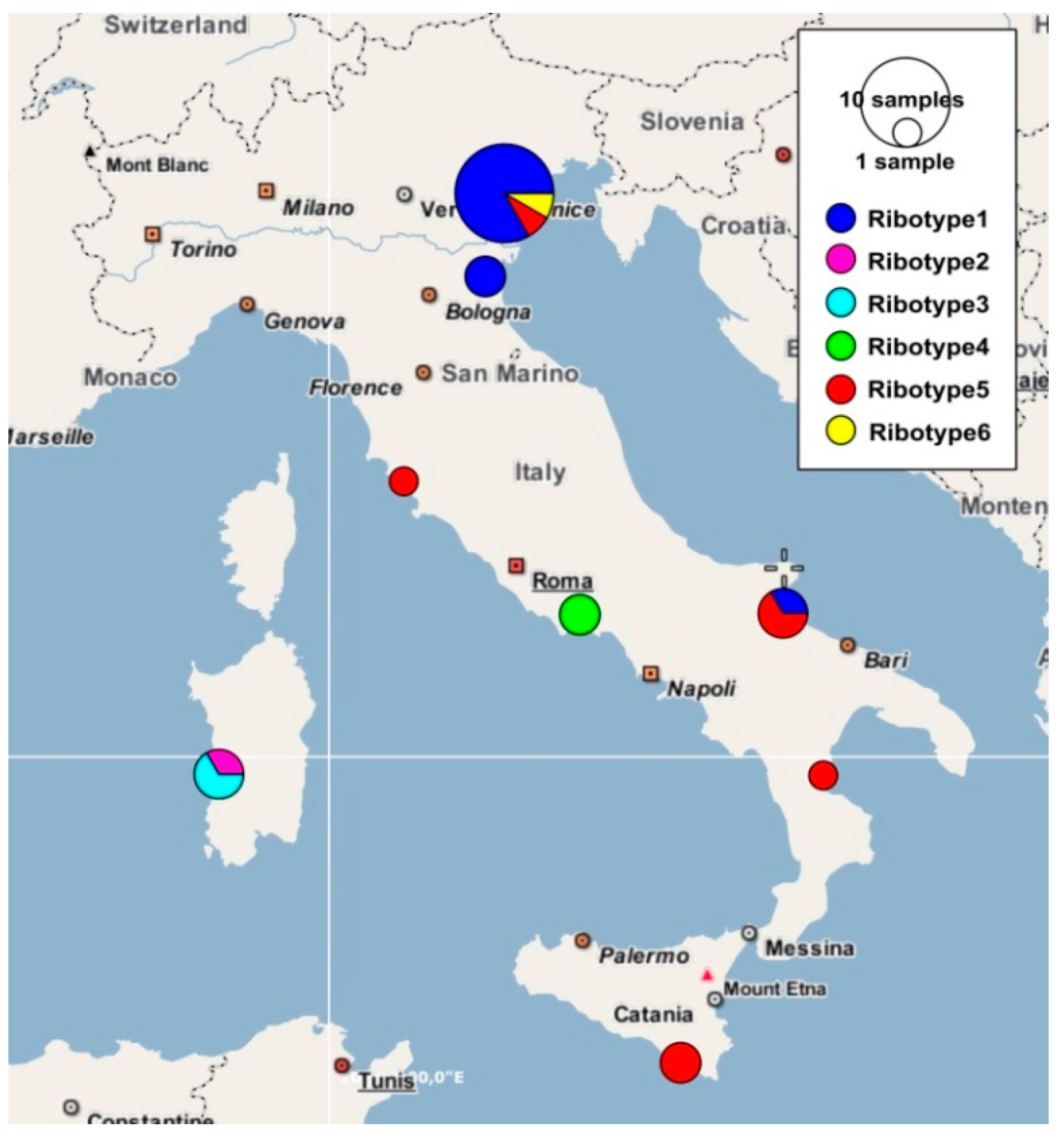
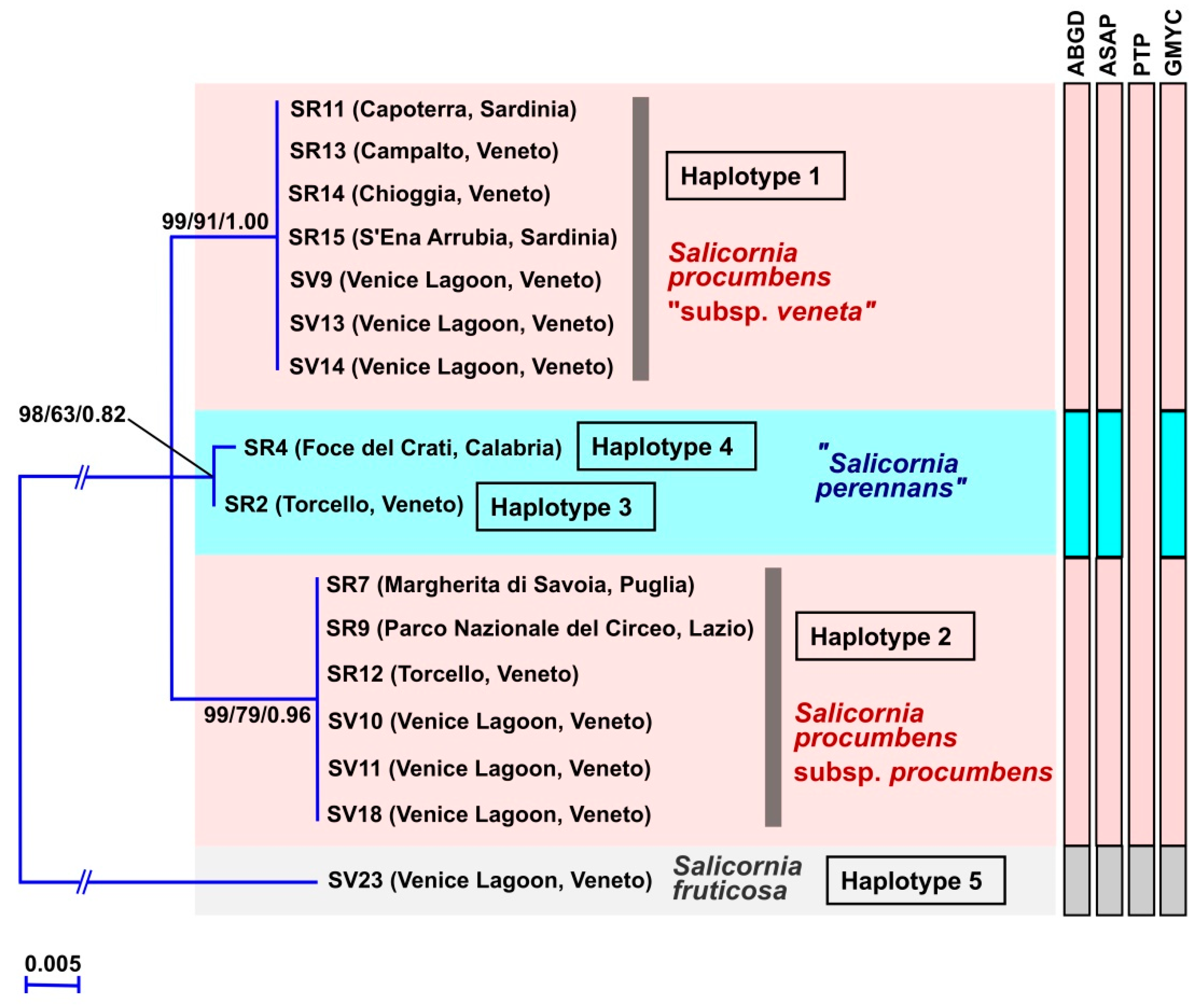
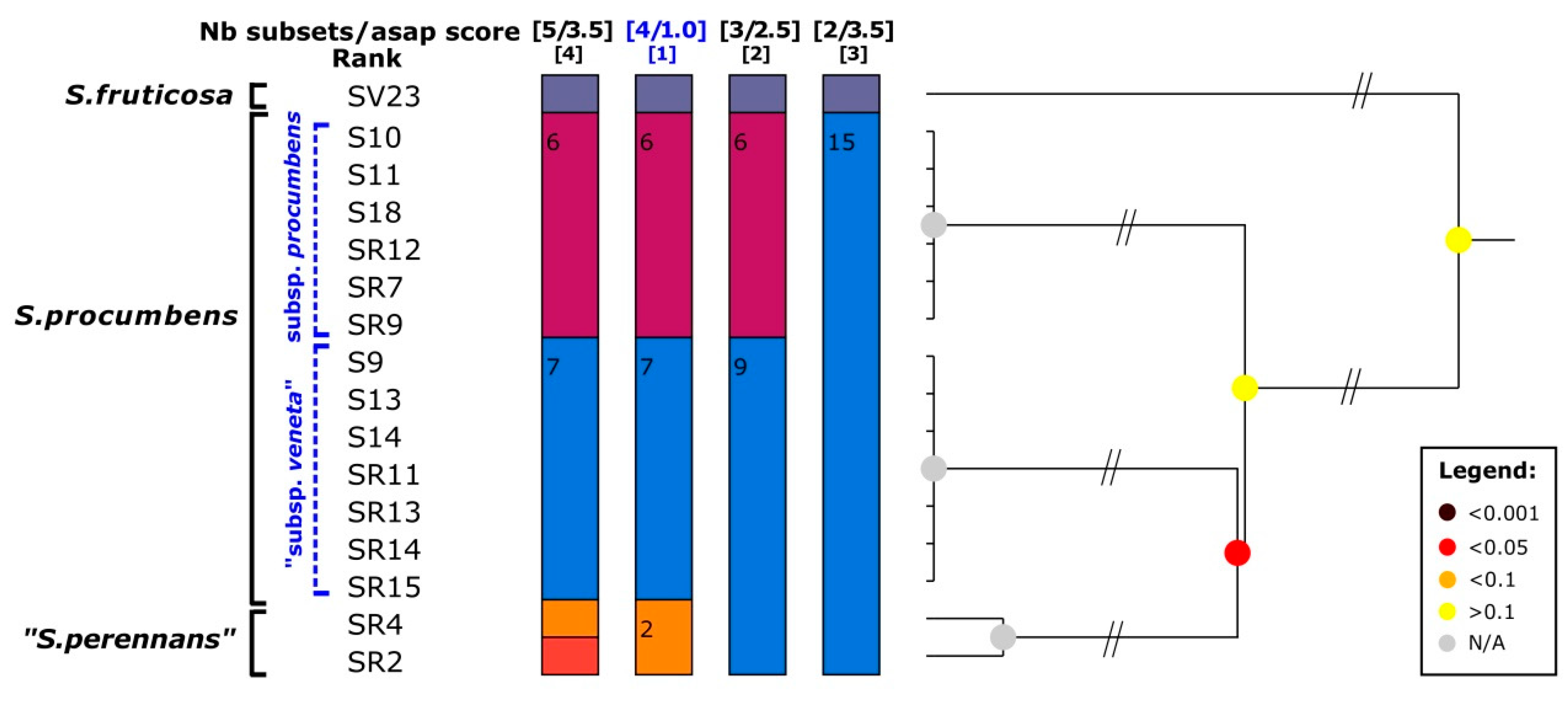
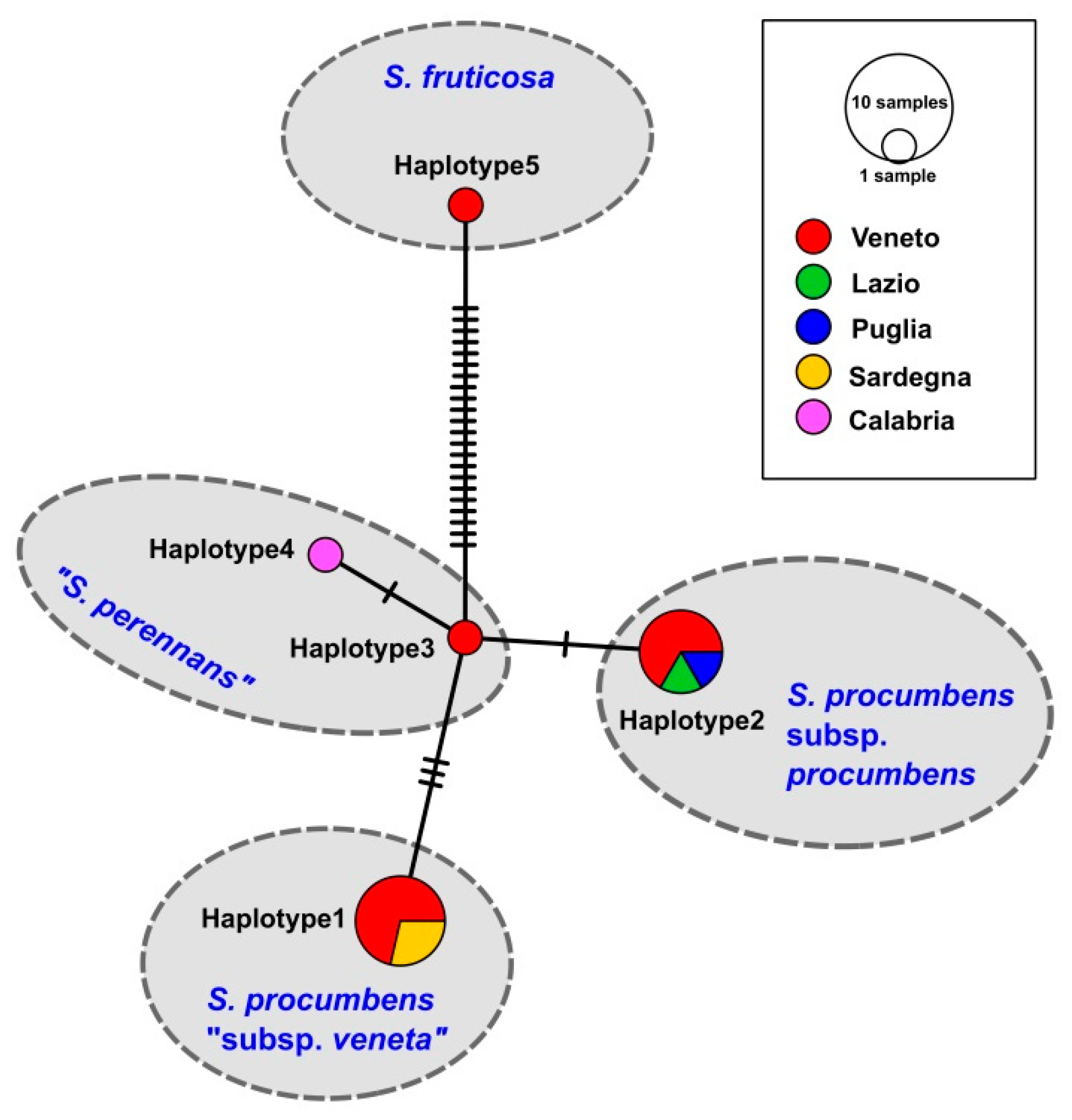
3. Results
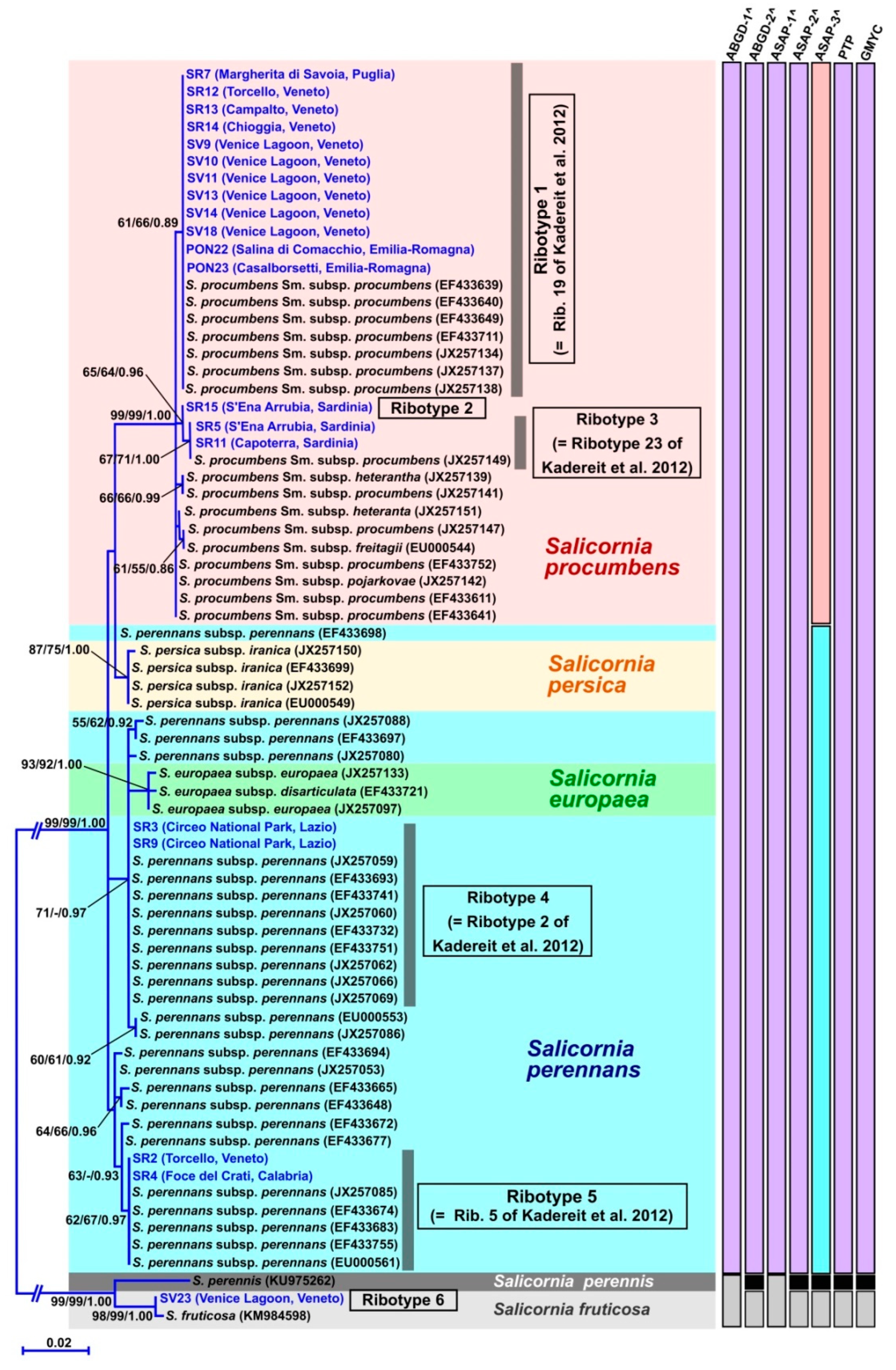
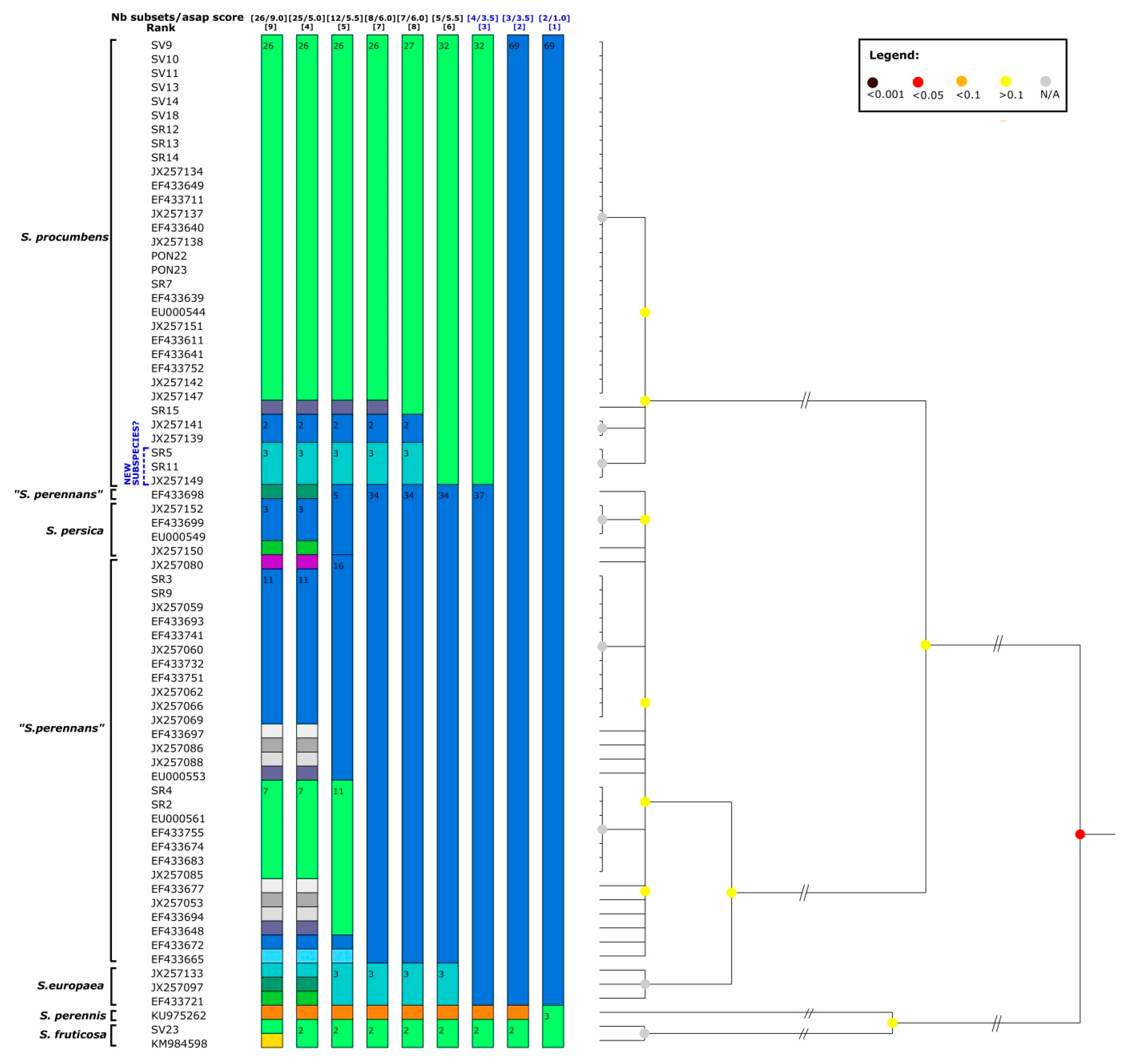
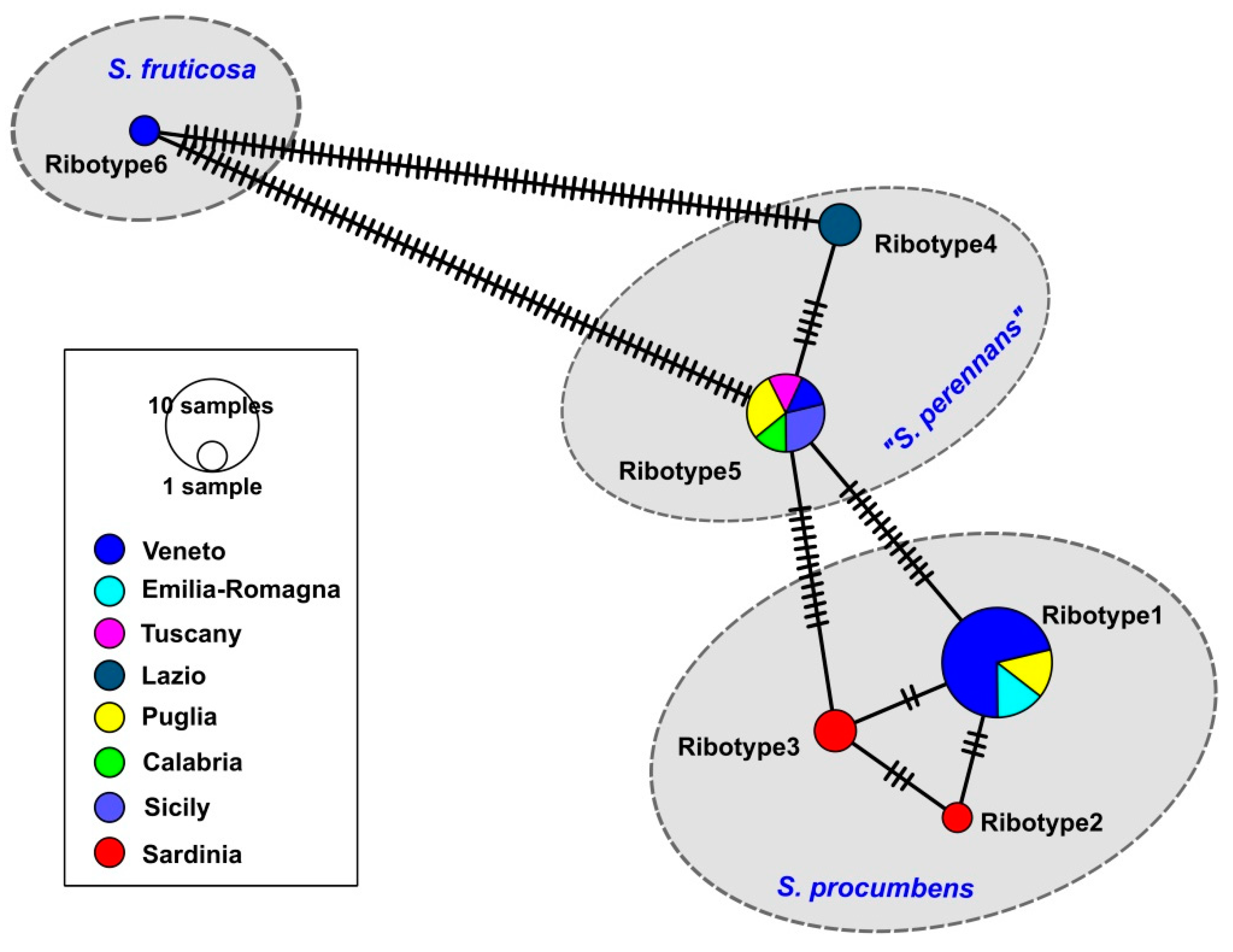
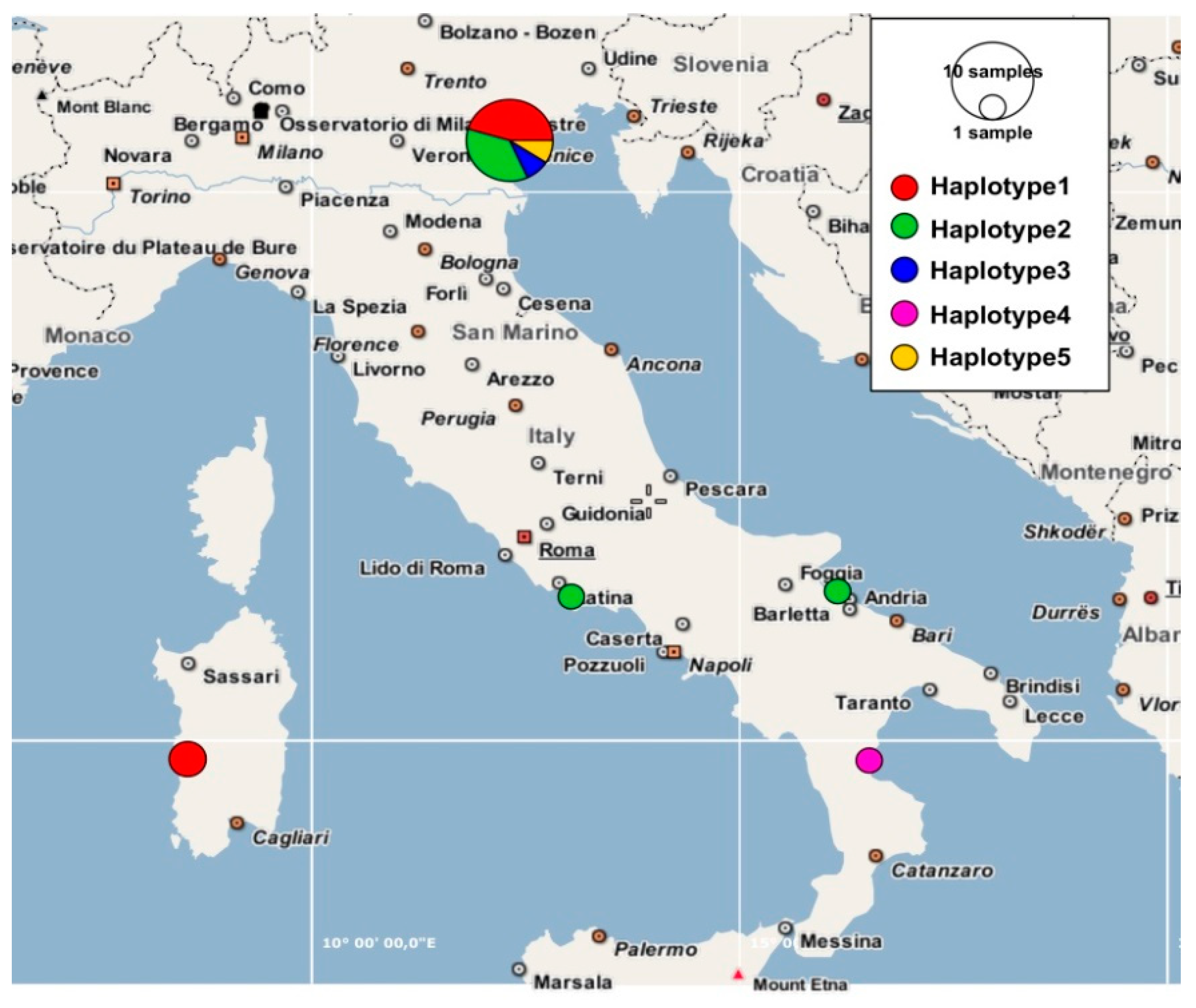
3.1. Molecular Study
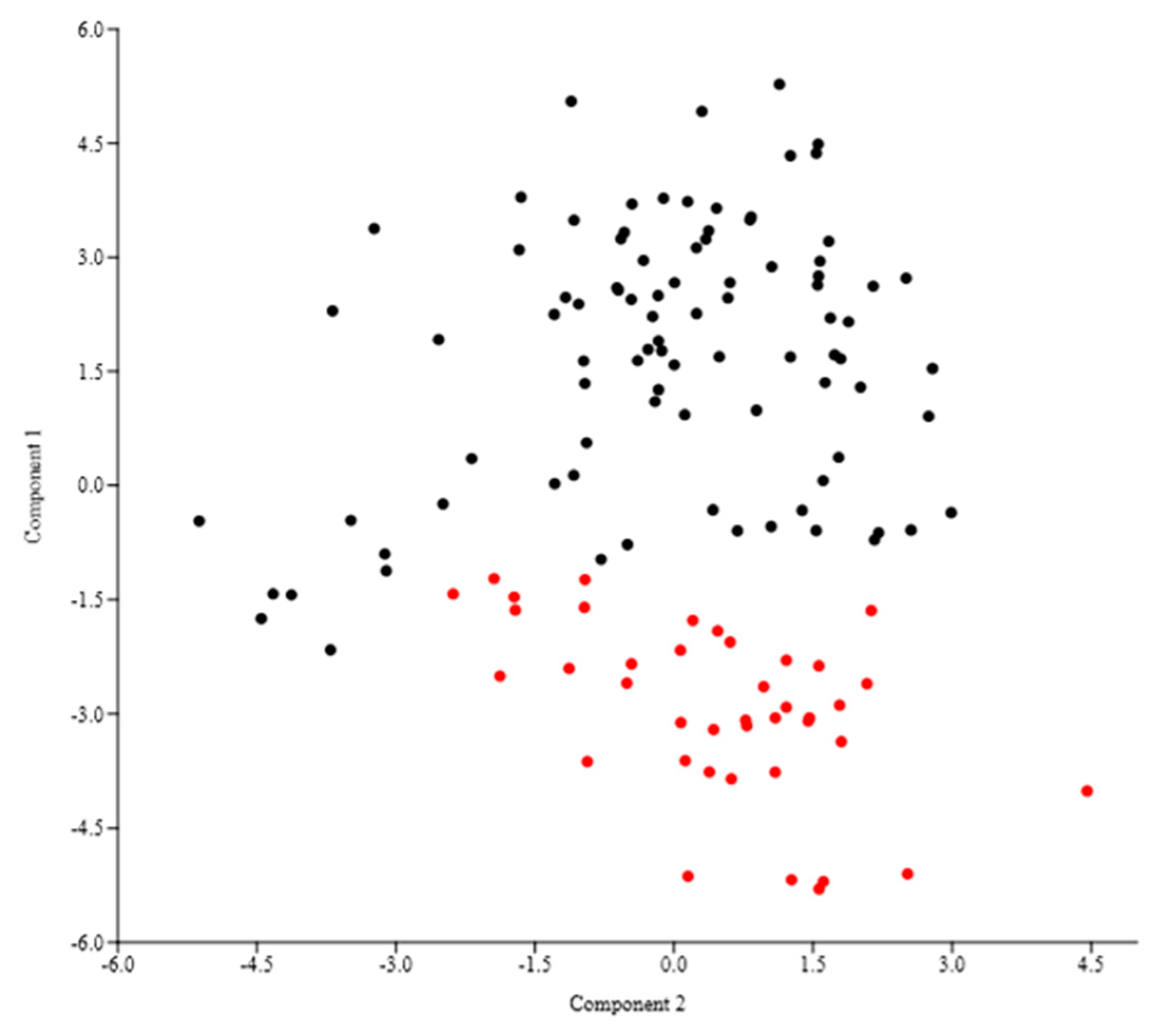
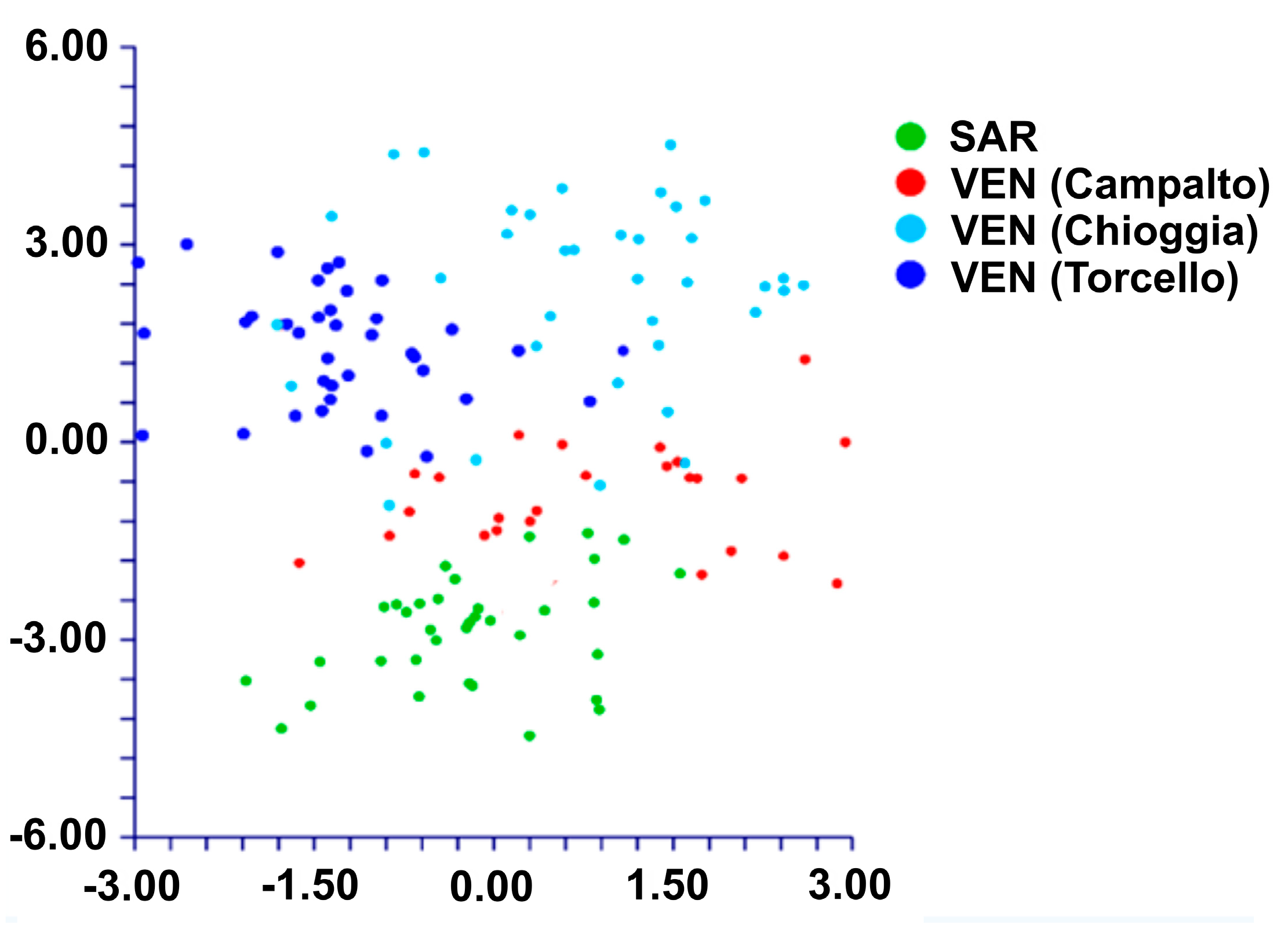
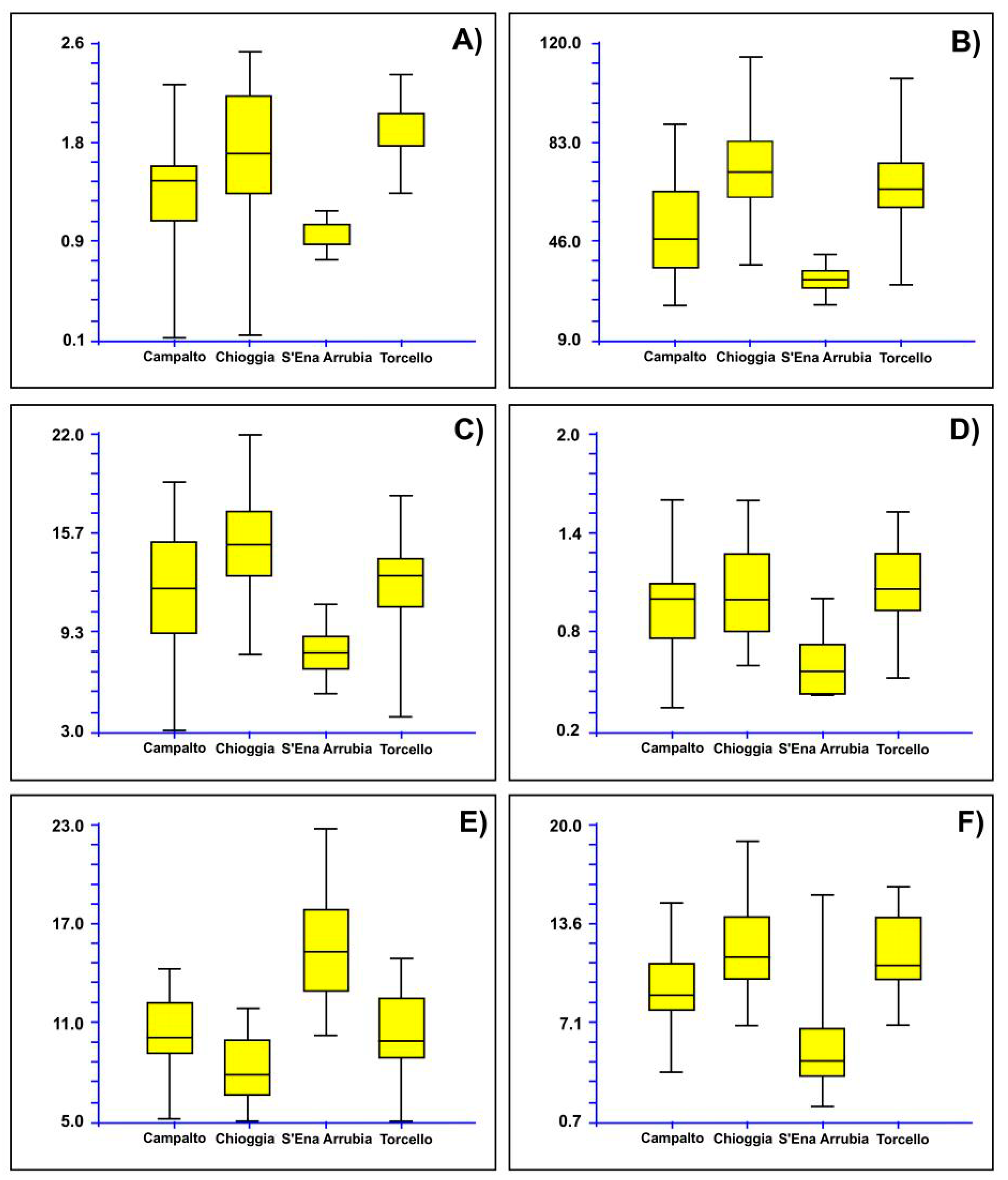
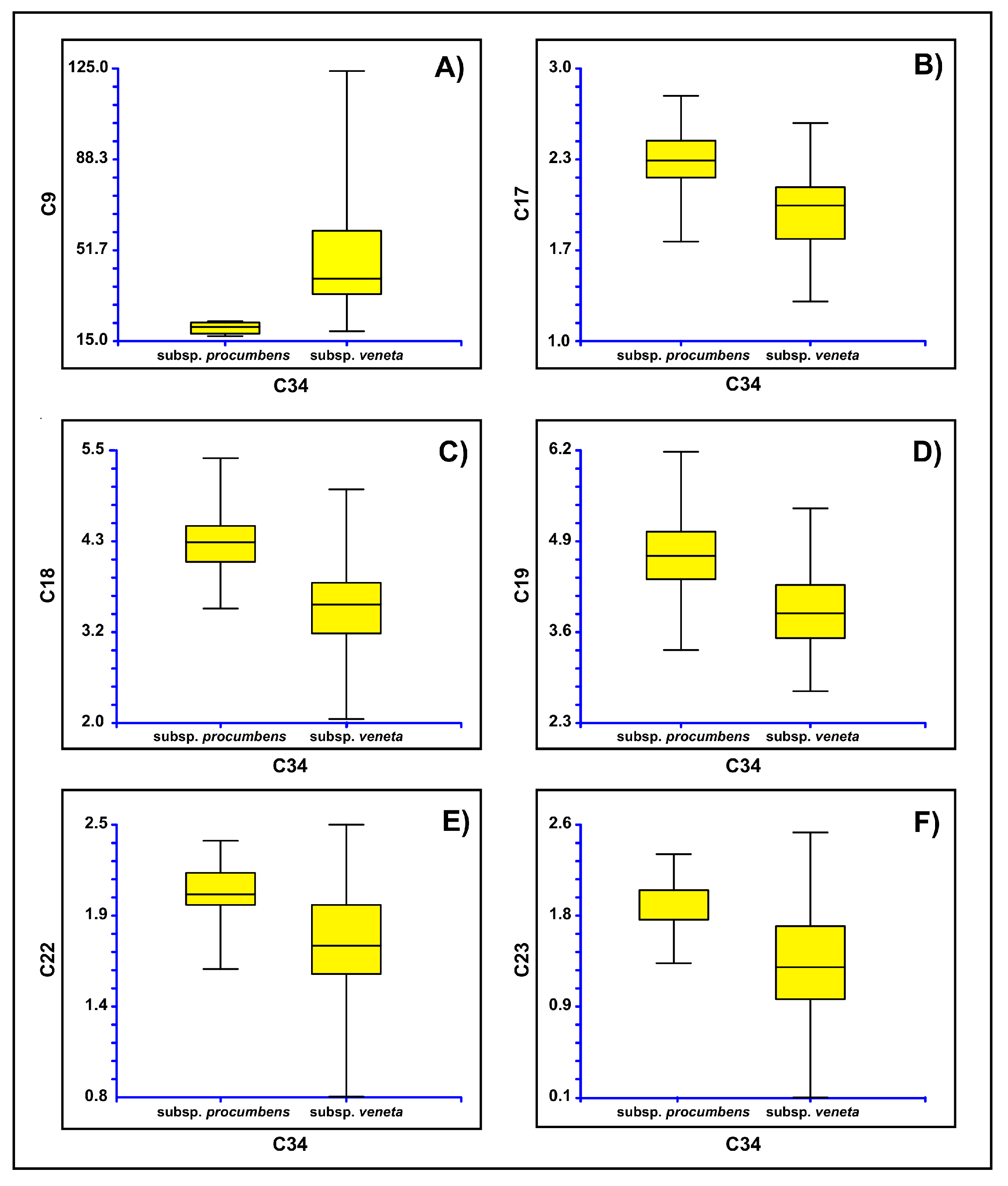
3.2. Morphometric Analyses
4. Discussion
5. Taxonomic Treatment
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanderpoorten, A.; Hardy, O.J.; Lambinon, J.; Raspé, O. Two reproductively isolated cytotypes and a swarm of highly inbred, disconnected populations: A glimpse into Salicornia’s evolutionary history and challenging taxonomy. J. Evol. Biol. 2011, 24, 630–644. [Google Scholar] [CrossRef]
- Hernández-Ledesma, P.; Berendsohn, W.G.; Borsch, T.; von Mering, S.; Akhani, H.; Arias, S.; Castañeda-Noa, I.; Eggli, U.; Eriksson, R.; Flores-Olvera, H.; et al. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales. Willdenowia 2015, 45, 281–383. [Google Scholar] [CrossRef] [Green Version]
- Fogli, S.; Brancaleoni, L.; Lambertini, C.; Gerdol, R. Mowing regime has different effects on reed stands in relation to habitat. J. Environ. Manag. 2014, 134, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iberite, M. Le salicornie: Metodologie di indagine e problematiche tassonomiche. Inform. Bot. Ital. 2004, 36, 508–511. [Google Scholar]
- Kadereit, G.; Mucina, L.; Freitag, H. Phylogeny of Salicornioideae (Chenopodiaceae): Diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 2006, 55, 617–642. [Google Scholar] [CrossRef]
- Kadereit, G.; Ball, P.; Beer, S.; Mucina, L.; Sokoloff, D.; Teege, P.; Yaprak, A.E.; Freitag, H. A taxonomic nightmare comes true: Phylogeny and biogeography of glassworts (Salicornia L. Chenopodiaceae). Taxon 2007, 56, 1143–1170. [Google Scholar] [CrossRef] [Green Version]
- Kadereit, G.; Piirainen, M.; Lambinon, J.; Vanderpoorten, A. Cryptic taxa should have names. Reflections on the glasswort genus Salicornia (Amaranthaceae). Taxon 2012, 61, 1227–1239. [Google Scholar] [CrossRef] [Green Version]
- Linnaeus, C. Species Plantarum; Laurentii Salvii: Holmiae, Sweden, 1753; Volume 1. [Google Scholar]
- Willdenow, C.L. Species Plantarum, 4th ed.; Impensis G. C. Nauk: Berlin, Germany, 1797; Volume 1. [Google Scholar]
- Smith, J.E. English Botany; or, Coloured Figures of British Plants with Their Essential Characters, Synonyms and Pances of Growth: To Which Will Be Added, Occasianal Remarks; J. Davis Chancery-Lane: London, UK, 1813; Volume 36. [Google Scholar]
- Akhani, H. Salicornia persica Akhani (Chenopodiaceae) a remarkable new species from Central Iran. Linz. Biol. Beitr. 2003, 35, 607–612. [Google Scholar]
- Akhani, H. Taxonomic revision of the genus Salicornia L. (Chenopodiaceae) in central and southern Iran. Pak. J. Bot. 2008, 40, 1635–1655. [Google Scholar]
- Piirainen, M.; Liebisch, O.; Kadereit, G. Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae)—A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 2017, 66, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Iberite, M.; Macario, M. Segnalazioni floristiche italiane. Salicornia dolichostachya Moss. (Chenopodiaceae). Specie nuova per l’Italia. Inform. Bot. Ital. 1992, 24, 201. [Google Scholar]
- Iberite, M. Salicornia L. In Flora d’Italia, 2nd ed.; Pignatti, S., Ed.; Ed agricole: Bologna, Italy, 2018; Volume 2, pp. 265–267. [Google Scholar]
- Moss, C.E. The International phytogeographical excursion in the British isles. Remarks on the characters and nomenclaturr of some critical plants noticed on the excursion. New Phytol. 1912, 11, 398–414. [Google Scholar] [CrossRef]
- Duval-Jouve, M.J. Des Salicornia de [‘Herault. Examen des caractereJ djfferefltiels et discussion de la s)’nonymie. B Soc. Bot. Fr. 1868, 15, 165–178. [Google Scholar] [CrossRef]
- Lausi, D. Descrizione di una nuova Salicornia dalla Laguna Veneta. Giorn. Bot. Ital. 1969, 103, 183–188. [Google Scholar] [CrossRef]
- Biondi, E.; Casavecchia, S. The halophilous retro-dune grasslands of the Italian Adriatic coastline. Braun-Blanquetia 2010, 46, 111–127. [Google Scholar]
- Bartolucci, F.; Domina, G.; Adorni, M.; Alessandrini, A.; Ardenghi, N.M.G.; Banfi, E.; Baragliu, G.A.; Bernardo, L.; Bertolli, A.; Biondi, E.; et al. Notulae to the Italian native vascular flora: 3. It. Bot. 2017, 3, 29–48. [Google Scholar]
- Roma-Marzio, F.; Peruzzi, L.; Bernanrdo, L.; Bartolucci, F.; De Ruvo, B.; De Ruvo, A.; Conti, F.; Giardini, M.; Domina, G.; Biondi, E.; et al. Nuove Segnalazioni Floristiche Italiane 3. Flora vascolare (010–021). Not. Della Soc. Bot. Ital. 2017, 1, 1–5. [Google Scholar]
- Yaprak, A.E.; Yurdakulol, E. Salicornia freitagii (Chenopodiaceae), a new species from Turkey. Ann. Bot. Fennici 2008, 45, 207–211. [Google Scholar] [CrossRef]
- Semenova, M.N. Salicornia pojarkovae sp. nov. In Flora Murmanskoi Oblasti; Gorodkov, B.N., Ed.; Akademia Nauk SSS: Moscow/Leningrad, Russia, 1956; Volume 3, p. 190. [Google Scholar]
- Beer, S.S.; Demina, O. A new species of Salicornia (Chenopodiaceae) from European Russia. Willdenowia 2005, 35, 253–257. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Cañadas, E.M.; Bacchetta, G. Disentangling the influence of environmental and anthropogenic factors on the distribution of endemic vascular plants in Sardinia. PLoS ONE 2017, 12, e0182539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fois, M.; Podda, L.; Médail, F.; Bacchetta, G. Endemic and alien vascular plant diversity in the small Mediterranean islands of Sardinia: Drivers and implications for their conservation. Biol. Conserv. 2020, 244, 108519. [Google Scholar] [CrossRef]
- Yao, P.C.; Gao, H.Y.; Wei, Y.N.; Zhang, J.H.; Chen, X.Y.; Li, H.-Q. Evaluating sampling strategy for DNA barcoding study of coastal and inland halo-tolerant Poaceae and Chenopodiaceae: A case study for increased sample size. PLoS ONE 2017, 12, e0185311. [Google Scholar] [CrossRef] [Green Version]
- Iberite, M. Contribution to the knowledge of the genus Salicornia L. (Chenopodiaceae) in Italy. Ann. Bot. 1996, 54, 145–154. [Google Scholar]
- Iberite, M.; Iamonico, D. Indagini sulla variabilità fenotipica di Salicornia veneta Pignatti & Lausi e S. emerici Duval–Jouve (Amaranthaceae). In PIPPs (Peripheral and Isolated Plant Population) ed Endemiti: Tassonomia, Filogenesi ed Evoluzione; Peruzzi, L., Bedini, G., Garbari, F., Eds.; Università di Pisa: Castiglioncello (Livorno), Italy, 2011; pp. 18–19. [Google Scholar]
- Iamonico, D.; Iberite, M. Amaranthaceae and Chenopodiaceae in Italy: Current understanding and future prospective. In Proceedings of the Symposium “Caryophyllales”: New Insights into the Phylogeny, Systematics and Morphological Evolution of the Order; Timonin, A.K., Sukhorukov, A.P., Harper, G.H., Nilova, M.V., Eds.; M. V. Lomonosov State University: Moscow, Russia, 2012; pp. 65–69. [Google Scholar]
- Iamonico, D.; Sukhorukov, A.P. Studies on the genus Atriplex (Chenopodiaceae) in Italy. VI. Names by Michele Tenore: Atriplex axillaris, A. diffusa, and A. polysperma. Hacquetia 2014, 13, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Freitag, H.; Iamonico, D. Lectotypification of the name Polycnemum majus (Amaranthaceae s.str.). Phytotaxa 2015, 202, 235–236. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Del Pino, I.; Iamonico, D. Jamesbondia, a new Subgenus of Alternanthera (Gomphrenoideae, Amaranthaceae) from Central America and the Caribbean Islands. Plant Biosyst. 2016, 150, 190–200. [Google Scholar] [CrossRef]
- Iamonico, D. Nomenclature survey of the genus Amaranthus (Amaranthaceae). 5. Moquin-Tandon’s names. Phytotaxa 2016, 273, 81–114. [Google Scholar] [CrossRef]
- Iamonico, D. Lectotypification of the Linnaean name Salsola sativa (Amaranthaceae s. lat.). Taxon 2016, 65, 872–874. [Google Scholar] [CrossRef]
- Iamonico, D. Remarks on the nomenclature of Betoideae (Chenopodiaceae/Amaranthaceae sensu APGIV). Phytotaxa 2019, 400, 145–164. [Google Scholar] [CrossRef]
- Iamonico, D. Nomenclature survey of the genus Amaranthus (Amaranthaceae s.s.). 8. About Amaranthus polygonoides s.l. and A. anderssonii, two related taxa described from the tropical regions of America with notes on their taxonomy. Acta Bot. Mex. 2020, 127, e1687. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/ih/ (accessed on 10 September 2022).
- Bruni, I.; De Mattia, F.; Martellos, S.; Galimberti, A.; Savadori, P.; Casiraghi, M.; Nimis, P.L.; Labra, M. DNA Barcoding as an Effective Tool in Improving a Digital Plant Identification System: A Case Study for the Area of Mt. Valerio, Trieste (NE Italy). PLoS ONE 2012, 7, e43256. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 2.71. 2009. Available online: http://mesquiteproject.org (accessed on 3 December 2022).
- Rambaut, A.; Drummond, A.J. Tracer, Version 1.5. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 3 December 2022).
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Topliss, J.G.; Costello, R.J. Chance correlations in structure-activity studies using multiple regression analysis. J. Med. Chem. 1972, 15, 1066–1068. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 81, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Stewart, S.; Ivy, M.I.; Anslyn, E.V. The use of principal component analysis and discriminant analysis in differential sensing routines. Chem. Soc. Rev. 2013, 43, 70–84. [Google Scholar] [CrossRef]
- Aristidis, L.; Vlassis, N.; Verbeek, J.J. The global k-means clustering algorithm. Pattern Recognit. 2003, 36, 451–461. [Google Scholar]
- Pelser, P.B.; Kennedy, A.H.; Tepe, E.J.; Shidler, J.B.; Nordenstam, B.; Kadereit, J.W.; Watson, L.E. Patterns and causes of incongruence between plastid and nuclear Senecioneae (Asteraceae) phylogenies. Am. J. Bot. 2010, 97, 856–873. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.G.; Larson, E.L. Hybridization, Introgression, and the Nature of Species Boundaries. J. Hered. 2014, 105, 795–809. [Google Scholar] [CrossRef]
- Štorchová, H.; Olson, M.S. The architecture of the chloroplast psbA-trnH non-coding region in angiosperms. Pl. Syst. Evol. 2007, 268, 235–256. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.-H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Mansion, G.; Rosenbaum, G.; Schoenenberger, N.; Bacchetta, G.; Rosselló, J.A.; Conti, E. Phylogenetic analysis informed by geological history supports multiple, sequential invasions of the Mediterranean Basin by the angiosperm family Araceae. Syst. Biol. 2008, 57, 269–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feliner, G.N. Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 265–278. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Locality and Specimens | ETS | psbA-trnH |
|---|---|---|---|
| SR2 | Italy, Veneto region, Venice province, Torcello islet, brackish hollows near the lagoon, 15 October 2005, M. Iberite | OM333820 | OP821351 |
| SR3 | Italy, Lazio region, Circeo National Park, Sabaudia, 6 October 2005, M. Iberite | OP821341 | - |
| SR4 | Italy, Calabria region, Cosenza province, protected area Foce del Crati, 1 November 2004, M. Iberite | OP821342 | OP821352 |
| SR5 | Italy, Sardinia region, Oristano province, S’Ena Arrubia, edge of the brackish pond, 31 October 2003, M. Iberite | OM333819 | - |
| SR7 | Italy, Puglia region, Barletta-Andria-Trani province, Margherita di Savoia, 28 August 2020, M. Iberite | OP821343 | OP821353 |
| SR9 | Italy, Lazio region, Circeo National Park, Sabaudia, 22 October 1994, M. Iberite | OP821344 | OP821354 |
| SR11 | Italy, Sardinia region, Cagliari province, Capoterra, 30 October 2003, M. Iberite | OM333821 | OP821355 |
| SR12 * | Italy, Veneto region, Venice province, Torcello islet, brackish hollows near the lagoon, 15 October 2005, M. Iberite (RO) | OM333822 | OP821356 |
| SR13 * | Italy, Veneto region, Venice province, Campalto, brackish hollows near the lagoon, 15 October 2005, M. Iberite (RO) | OM333823 | OP821357 |
| SR14 * | Italy, Veneto region, Venice province, Chioggia, brackish hollows near the lagoon, 16 October 2005, M. Iberite (RO) | OM333824 | OP821358 |
| SR15 * | Italy, Sardinia region, Oristano province, S’Ena Arrubia, edge of the brackish pond, 30 September 2012, C. Pepe (RO) | OM333825 | OP821359 |
| SV9 | Italy, Veneto region, Venice province, North Lagoon, 26 June 2018, A. Sfriso | OP821345 | OP821360 |
| SV10 | Italy, Veneto region, Venice province, North Lagoon, 26 June 2018, A. Sfriso | OP821346 | OP821361 |
| SV11 | Italy, Veneto region, Venice province, North Lagoon, 26 June 2018, A. Sfriso | OP821347 | OP821362 |
| SV13 | Italy, Veneto region, Venice province, North Lagoon, 10 September 2018, A. Sfriso | OP821348 | OP821363 |
| SV14 | Italy, Veneto region, Venice province, North Lagoon, 10 September 2018, A. Sfriso | OP821349 | OP821364 |
| SV18 | Italy, Veneto region, Venice province, North Lagoon, 27 September 2018, A. Sfriso | OP821350 | OP821365 |
| SV23 | Italy, Veneto region, Venice province, North Lagoon, 10 October 2019, A. Sfriso | OM333826 | OP821366 |
| PON22 | Italy, Emilia-Romagna region, Ferrara province, Salina di Comacchio, 10 August 2022, L. Brancaleoni | OP328927 | - |
| PON23 | Italy, Emilia-Romagna region, Ravenna province, Casalborsetti, 10 August 2022, L. Brancaleoni | OP328928 | - |
| General Morphology | 1. height of the plant (mm) |
| 2. height of the plant from rooting point to the first branching point (mm) | |
| 3. number of internodes | |
| 4. length of the first internode (mm) | |
| 5. length of the second internode (mm) | |
| 6. length of the penultimate internode (mm) | |
| 7. length of the ultimate internode (mm) | |
| 8. length of the longest first primary branch (mm) | |
| 9. length of the ultimate branch (mm) | |
| 10. maximum number of secondaries on a primary branch | |
| 11. length of longest secondary branch (mm) | |
| 12. length of longest tertiary branch (mm) | |
| Characters of the Terminal Spike | 13. spike length (mm) |
| 14. number of fertile segments | |
| 15. number of sterile segments | |
| 16. maximum width of the third fertile segment (mm) | |
| 17. width of the middle flower (the third segment) (mm) | |
| 18. width of the three flowers (the third segment) (mm) | |
| 19. minimum width of the second fertile segment (mm) | |
| 20. maximum width of the second fertile segment (mm) | |
| 21. distance from the tip of the third fertile segments to apex of middle flower (mm) | |
| 22. height of the middle flower of the third segment (mm) | |
| 23. height of the side flower of the third segment (mm) | |
| 24. height of the scarious margin of the second fertile segment (mm) | |
| 25. color of the sterile segments (green or yellow, pinkish, reddish, green with red margin) * | |
| 26. color of the flowers of the third segment (green or yellow, pinkish, reddish, green with red margin) * | |
| 27. color of the fertile segments (green or yellow, pinkish, reddish, green with red margin) * | |
| 28. distribution of coloration (basal or even, apical) * |
| Test Statistic | Test Value | F-Ratio | p (0.05) |
|---|---|---|---|
| Wilks’ Lambda | 0.170952 | 18.88 | 0.0000001 |
| Hotelling–Lawley Trace | 4.849603 | 18.88 | 0.0000001 |
| Pillai’s Trace | 0.829048 | 18.88 | 0.0000001 |
| Roy’s Largest Root | 4.849603 | 18.88 | 0.0000001 |
| No. of Clusters | Component | Between Mean Square | Within Mean Square | F-Ratio |
|---|---|---|---|---|
| 2 | 1 | 87.83528 | 0.3335492 | 263.34 |
| 2 | 18.58399 | 0.9203334 | 20.19 | |
| 3 | 1.327774 | 1.081567 | 1.23 | |
| 4 | 4.31783 | 1.056084 | 4.09 | |
| 5 | 1.842877 | 1.126392 | 1.64 | |
| 3 | 1 | 45.3238 | 0.3151879 | 143.80 |
| 2 | 13.40127 | 0.8662724 | 15.47 | |
| 3 | 30.0381 | 0.6544046 | 45.90 | |
| 4 | 8.941694 | 0.9634211 | 9.28 | |
| 5 | 2.232657 | 1.11531 | 2.00 | |
| 4 | 1 | 31.93618 | 0.2790256 | 114.46 |
| 2 | 30.4127 | 0.3918748 | 77.61 | |
| 3 | 23.55327 | 0.580306 | 40.59 | |
| 4 | 8.110217 | 0.9224969 | 8.79 | |
| 5 | 0.5629421 | 1.144354 | 0.49 | |
| 5 | 1 | 21.64586 | 0.3504851 | 61.76 |
| 2 | 25.13895 | 0.3247632 | 77.41 | |
| 3 | 17.15698 | 0.5999466 | 28.60 | |
| 4 | 14.80934 | 0.6669765 | 22.20 | |
| 5 | 12.01765 | 0.8042226 | 14.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciuto, K.; Wolf, M.A.; Sfriso, A.; Brancaleoni, L.; Iberite, M.; Iamonico, D. Molecular and Morphometric Update on Italian Salicornia (Chenopodiaceae), with a Focus on the Species S. procumbens s. l. Plants 2023, 12, 375. https://doi.org/10.3390/plants12020375
Sciuto K, Wolf MA, Sfriso A, Brancaleoni L, Iberite M, Iamonico D. Molecular and Morphometric Update on Italian Salicornia (Chenopodiaceae), with a Focus on the Species S. procumbens s. l. Plants. 2023; 12(2):375. https://doi.org/10.3390/plants12020375
Chicago/Turabian StyleSciuto, Katia, Marion A. Wolf, Adriano Sfriso, Lisa Brancaleoni, Mauro Iberite, and Duilio Iamonico. 2023. "Molecular and Morphometric Update on Italian Salicornia (Chenopodiaceae), with a Focus on the Species S. procumbens s. l." Plants 12, no. 2: 375. https://doi.org/10.3390/plants12020375