
Testing a Model of Pacific Oysters’ (Crassostrea gigas) Growth in the Adriatic Sea: Implications for Aquaculture Spatial Planning

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Model Parameter Estimation
2.2. Mapping Indicators
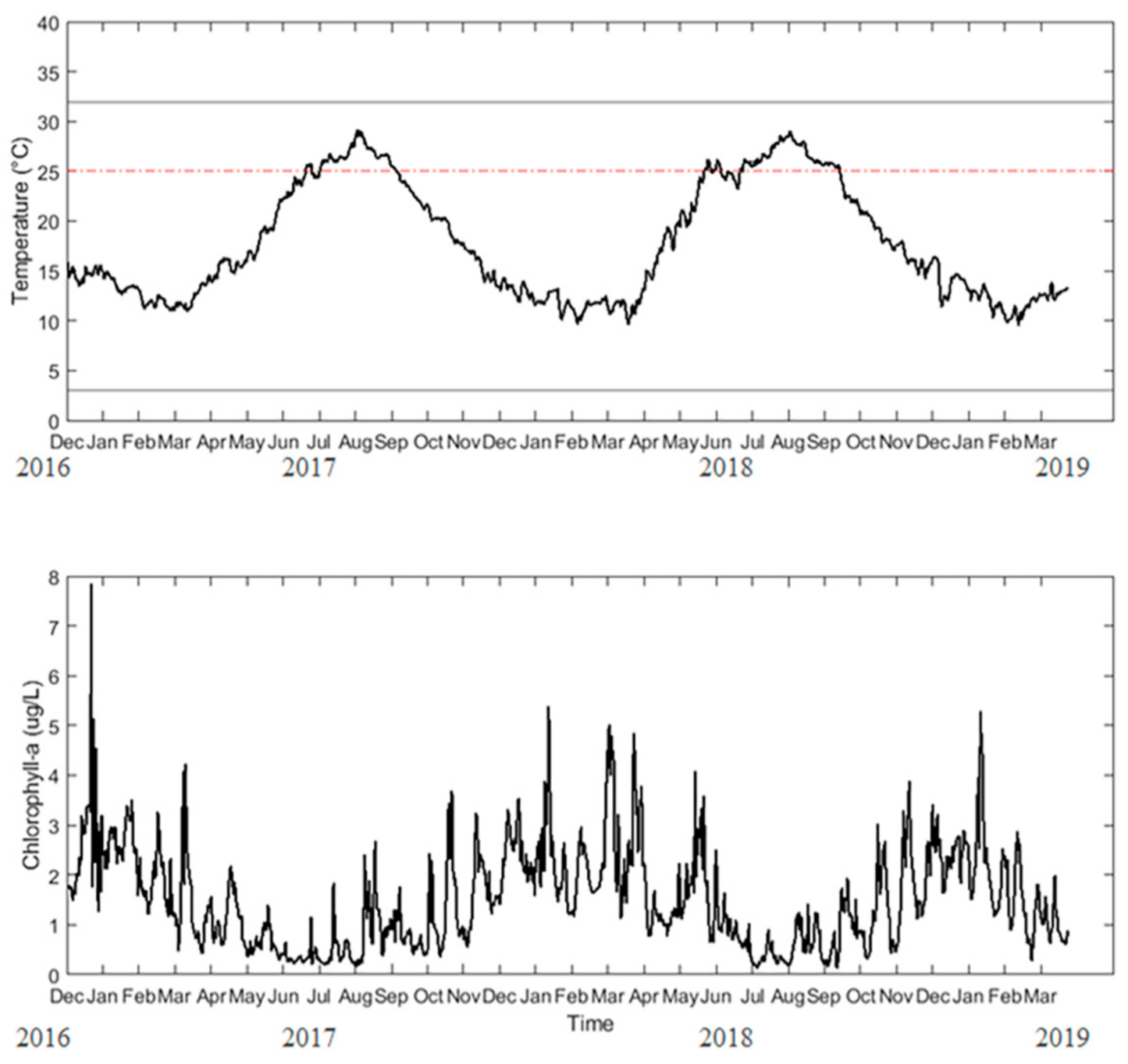
2.3. In Situ Data Collection
2.4. Forcing Functions
3. Results
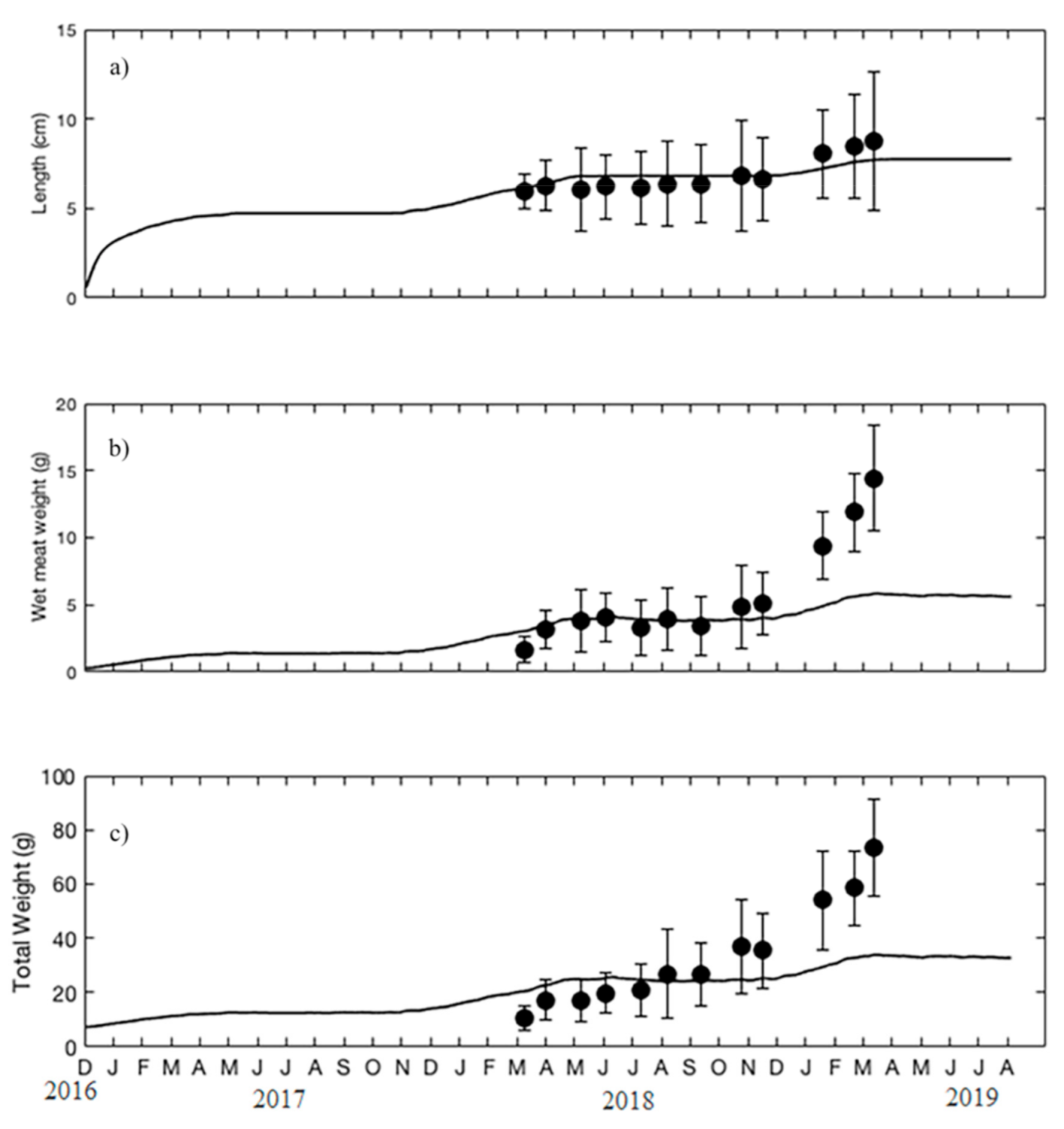
3.1. Parameter Estimation
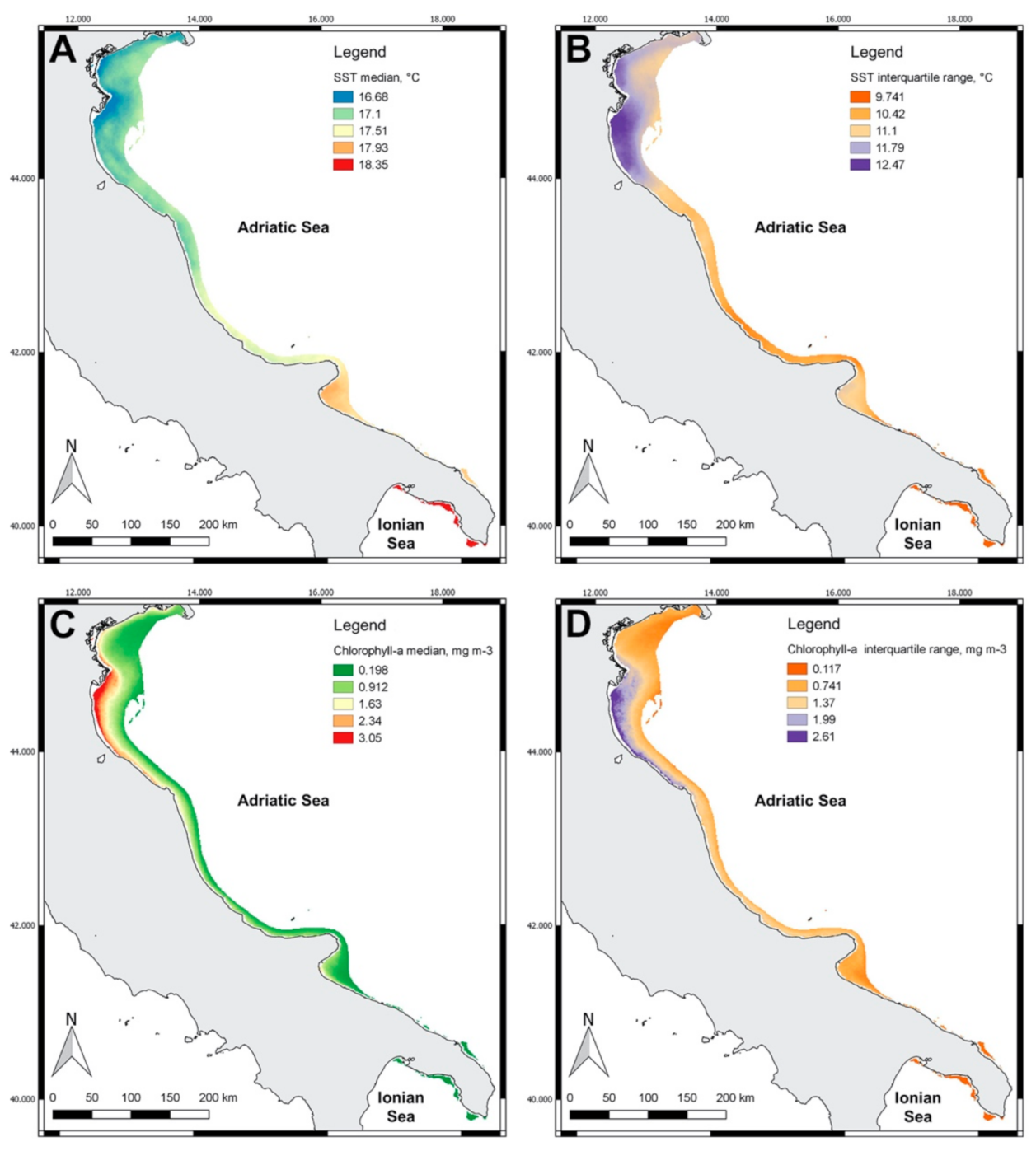
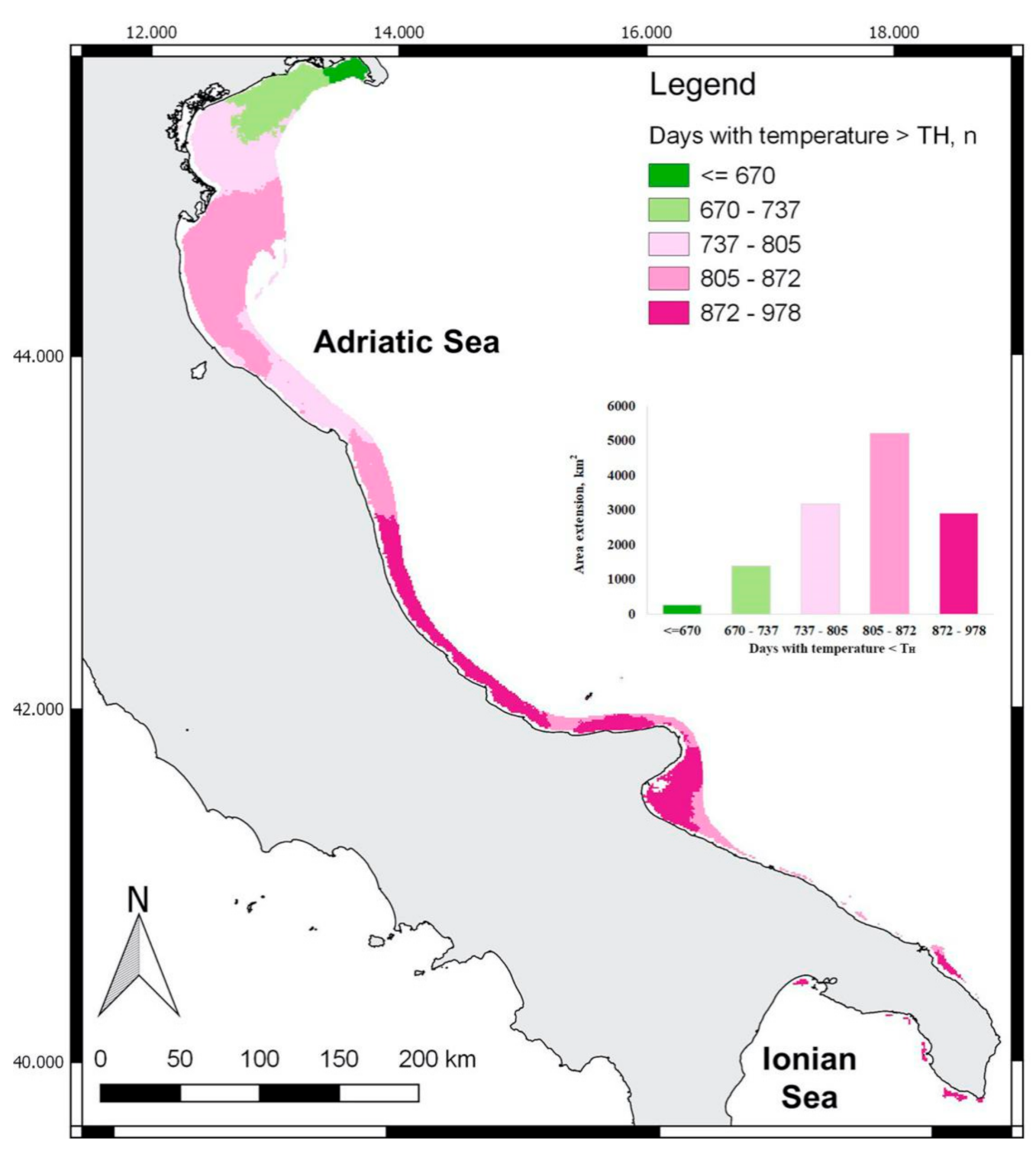
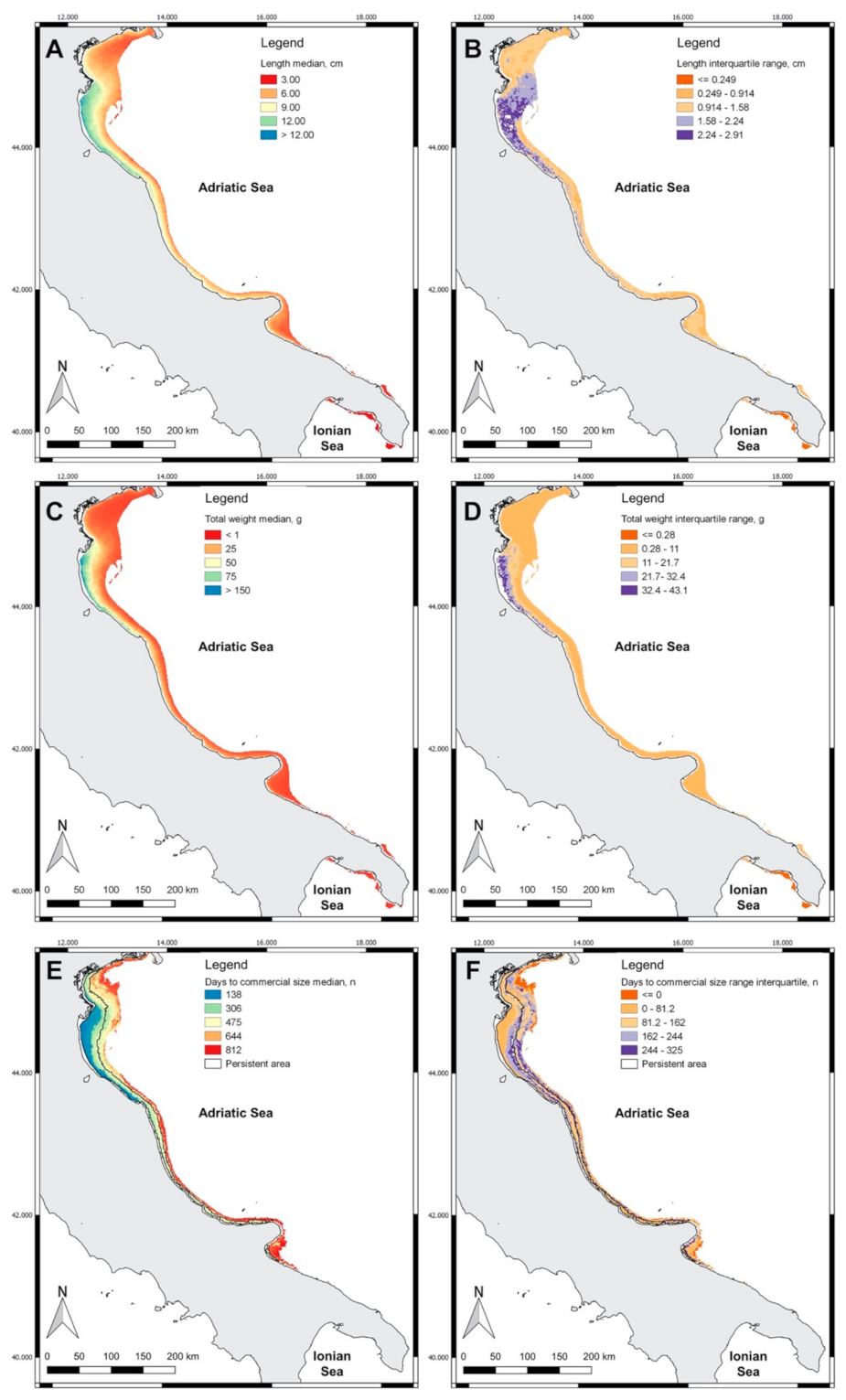
3.2. Spatial Distribution of Indicators
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Number before Sampling | Oysters Taken | Mortalities | Number after Sampling | Number in Each Basket (Average) | Volume of Water per Oyster (L) |
|---|---|---|---|---|---|---|
| March | 864 | 60 | 804 | 201 | 0.119 | |
| April | 804 | 60 | 744 | 186 | 0.129 | |
| May | 744 | 60 | 684 | 171 | 0.140 | |
| June | 684 | 60 | 624 | 156 | 0.153 | |
| July | 624 | 60 | 2 | 562 | 140.5 | 0.170 |
| August | 562 | 40 | 20 | 502 | 125.5 | 0.191 |
| September | 502 | 40 | 67 | 395 | 98.75 | 0.243 |
| October | 395 | 40 | 9 | 346 | 86.5 | 0.277 |
| December | 346 | 40 | 306 | 76.5 | 0.313 | |
| January | 306 | 40 | 266 | 66.5 | 0.360 | |
| February | 266 | 40 | 226 | 56.5 | 0.424 |
References
- Subasinghe, R.; Soto, D.; Jia, J. Global aquaculture and its role in sustainable development. Rev. Aquac. 2009, 1, 2–9. [Google Scholar] [CrossRef]
- Merino, G.; Barange, M.; Blanchard, J.L.; Harle, J.; Holmes, R.; Allen, I.; Allison, E.H.; Badjeck, M.C.; Dulvy, N.K.; Holt, J.; et al. Can marine fisheries and aquaculture meet fish demand from a growing human population in a changing climate? Glob. Environ. Chang. 2012, 22, 795–806. [Google Scholar] [CrossRef]
- Brummett, R. Agriculture and Environmental Services Department Growing Aquaculture in Sustainable Ecosystems. Available online: http://documents1.worldbank.org/curated/en/556181468331788600/pdf/788230BRI0AES00without0the0abstract.pdf (accessed on 16 March 2021).
- Shumway, S.E.; Davis, C.; Downey, R.; Karney, R.; Kraeuter, J.; Parsons, J.; Rheault, R.; Wikfors, G. Shellfish aquaculture—In praise of sustainable economies and environments. World Aquac. 2003, 34, 8–10. [Google Scholar]
- Cranford, P.; Dowd, M.; Grant, J.; Hargrave, B.; Mcgladdery, S. Ecosystem Level Effects of Marine Bivalve Aquaculture. Available online: https://waves-vagues.dfo-mpo.gc.ca/Library/365644.pdf (accessed on 16 March 2021).
- Wijsman, J.W.M.; Troost, K.; Fang, J.; Roncarati, A. Global production of marine bivalves. Trends and challenges. In Goods and Services of Marine Bivalves; Springer: Berlin, Germany, 2019; ISBN 9783319967769. [Google Scholar]
- Froehlich, H.E.; Gentry, R.R.; Halpern, B.S. Conservation aquaculture: Shifting the narrative and paradigm of aquaculture’s role in resource management. Biol. Conserv. 2017, 215, 162–168. [Google Scholar] [CrossRef]
- Smaal, A.C.; Ferreira, J.G.; Grant, J.; Petersen, J.K.; Strand, Ø. Goods and Services of Marine Bivalves; Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-319-96775-2. [Google Scholar]
- Lovelock, C.E.; Duarte, C.M. Dimensions of Blue Carbon and emerging perspectives. Biol. Lett. 2019, 15, 20180781. [Google Scholar] [CrossRef]
- Galparsoro, I.; Murillas, A.; Pinarbasi, K.; Sequeira, A.M.M.; Stelzenmüller, V.; Borja, Á.; O’Hagan, A.M.; Boyd, A.; Bricker, S.; Garmendia, J.M.; et al. Global stakeholder vision for ecosystem-based marine aquaculture expansion from coastal to offshore areas. Rev. Aquac. 2020, 12, 2061–2079. [Google Scholar] [CrossRef]
- Food and Agricultural Organisation (FAO). Cultured Aquatic Species Information Programme (Oreochromis niloticus); FAO: Rome, Italy, 2015. [Google Scholar]
- King, N.G.; Wilmes, S.B.; Smyth, D.; Tinker, J.; Robins, P.E.; Thorpe, J.; Jones, L.; Malham, S.K. Climate change accelerates range expansion of the invasive non-native species, the Pacific oyster, Crassostrea gigas. ICES J. Mar. Sci. 2020. [Google Scholar] [CrossRef]
- Massa, F.; Onofri, L.; Fezzardi, D. Aquaculture in the Mediterranean and the Black Sea: A Blue Growth perspective. Handb. Econ. Manag. Sustain. Ocean. 2017, 93–123. [Google Scholar] [CrossRef]
- Roncarati, A.; Felici, A.; Magi, G.E.; Bilandžić, N.; Melotti, P. Growth and survival of cupped oysters (Crassostrea gigas) during nursery and pregrowing stages in open sea facilities using different stocking densities. Aquac. Int. 2017, 25, 1777–1785. [Google Scholar] [CrossRef]
- Palmer, S.C.J.; Gernez, P.M.; Thomas, Y.; Simis, S.; Miller, P.I.; Glize, P.; Barillé, L. Remote Sensing-Driven Pacific Oyster (Crassostrea gigas) Growth Modeling to Inform Offshore Aquaculture Site Selection. Front. Mar. Sci. 2020, 6, 802. [Google Scholar] [CrossRef]
- Thomas, Y.; Pouvreau, S.; Alunno-Bruscia, M.; Barillé, L.; Gohin, F.; Bryère, P.; Gernez, P. Global change and climate-driven invasion of the Pacific oyster (Crassostrea gigas) along European coasts: A bioenergetics modelling approach. J. Biogeogr. 2016, 43, 568–579. [Google Scholar] [CrossRef] [Green Version]
- Coppini, G.; Lyubarstev, V.; Pinardi, N.; Colella, S.; Santoleri, R.; Christiansen, T. Chl a trends in European seas estimated using ocean-colour products. Ocean Sci. Discuss. 2012, 9, 1481–1518. [Google Scholar] [CrossRef] [Green Version]
- Pastor, F.; Valiente, J.A.; Palau, J.L. Sea Surface Temperature in the Mediterranean: Trends and Spatial Patterns (1982–2016). Pure Appl. Geophys. 2018, 175, 4017–4029. [Google Scholar] [CrossRef] [Green Version]
- Bougrier, S.; Geairon, P.; Deslous-Paoli, J.M.; Bacher, C.; Jonquières, G. Allometric relationships and effects of temperature on clearance and oxygen consumption rates of Crassostrea gigas (Thunberg). Aquaculture 1995, 134, 143–154. [Google Scholar] [CrossRef]
- Jossart, J.; Theuerkauf, S.J.; Wickliffe, L.C.; Morris, J.A., Jr. A. Applications of Spatial Autocorrelation Analyses for Marine Aquaculture Siting. Front. Mar. Sci. 2020, 6, 806. [Google Scholar] [CrossRef]
- Porporato, E.M.D.; Pastres, R.; Brigolin, D. Site Suitability for Finfish Marine Aquaculture in the Central Mediterranean Sea. Front. Mar. Sci. 2020, 6, 772. [Google Scholar] [CrossRef] [Green Version]
- Kooijman, S.A.L.M. Dynamic Energy Budget Theory for Metabolic Organisation, 3rd ed.; Cambridge University Press: Cambridge, UK, 2009; ISBN 9780511805400. [Google Scholar]
- Bacher, C.; Gangnery, A. Use of dynamic energy budget and individual based models to simulate the dynamics of cultivated oyster populations. J. Sea Res. 2006, 56, 140–155. [Google Scholar] [CrossRef] [Green Version]
- Mangano, M.C.; Giacoletti, A.; Sarà, G. Dynamic Energy Budget provides mechanistic derived quantities to implement the ecosystem based management approach. J. Sea Res. 2019, 143, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Alunno-Bruscia, M.; Bourlès, Y.; Maurerb, D.; Robertc, S.; Mazuriéd, J.; Gangnerye, A.; Goulletquerf, P.; Pouvreaua, S. A single bio-energetics growth and reproduction model for the oyster. J. Sea Res. 2011, 66, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Pouvreau, S.; Bourles, Y.; Lefebvre, S.; Gangnery, A.; Alunno-Bruscia, M. Application of a dynamic energy budget model to the Pacific oyster, Crassostrea gigas, reared under various environmental conditions. J. Sea Res. 2006, 56, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Sarà, G.; Reid, G.K.; Rinaldi, A.; Palmeri, V.; Troell, M.; Kooijman, S.A.L.M. Growth and reproductive simulation of candidate shellfish species at fish cages in the Southern Mediterranean: Dynamic Energy Budget (DEB) modelling for integrated multi-trophic aquaculture. Aquaculture 2012, 324–325, 259–266. [Google Scholar] [CrossRef]
- Brigolin, D.; Porporato, E.M.D.; Prioli, G.; Pastres, R. Making space for shellfish farming along the Adriatic coast. ICES J. Mar. Sci. 2017, 74, 1540–1551. [Google Scholar] [CrossRef] [Green Version]
- Troost, T.A.; Wijsman, J.W.M.; Saraiva, S.; Freitas, V. Modelling shellfish growth with dynamic energy budget models: An application for cockles and mussels in the Oosterschelde (southwest Netherlands). Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3567–3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, C.; Hendriks, A.J. Half-saturation constants in functional responses. Glob. Ecol. Conserv. 2014, 2, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Bourlès, Y.; Pouvreau, S.; Tollu, G.; Leguay, D.; Arnaud, D.; Goulletquer, P.; Kooijman, S. Modelling growth and reproduction of the Pacific oyster Crassostrea gigas: Advances in the oyster-DEB model through application to a coastal pond. J. Sea Res. 2009, 62, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.S.; Ross, A.H.; Schiel, D.R. Functional descriptions of feeding and energetics of the Pacific oyster Crassostrea gigas in New Zealand. Mar. Ecol. Prog. Ser. 2000, 208, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.S.; Schiel, D.R. A dynamic energy budget model: Parameterisation and application to the Pacific oyster Crassostrea gigas in New Zealand waters. J. Exp. Mar. Biol. Ecol. 2008, 361, 42–48. [Google Scholar] [CrossRef]
- Dixon, P.M. Bootstrap Resampling. In Encyclopedia of Environmetrics; John Wiley & Sons, Ltd.: Chichester, UK, 2006. [Google Scholar]
- van der Veer, H.W.; Cardoso, J.F.M.F.; van der Meer, J. The estimation of DEB parameters for various Northeast Atlantic bivalve species. J. Sea Res. 2006, 56, 107–124. [Google Scholar] [CrossRef]
- Deslous-Paoli, J.-M.; Héral, M. Biochemical composition and energy value of Crassostrea gigas (Thunberg) cultured in the bay of Marennes-Oléron. Aquat. Living Resour. 1988, 1, 239–249. [Google Scholar] [CrossRef]
- Rosland, R.; Strand, Ø.; Alunno-Bruscia, M.; Bacher, C.; Strohmeier, T. Applying Dynamic Energy Budget (DEB) theory to simulate growth and bio-energetics of blue mussels under low seston conditions. J. Sea Res. 2009, 62, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Cilenti, L.; Scirocco, T.; Specchiulli, A.; Vitelli, M.L.; Manzo, C.; Fabbrocini, A.; Santucci, A.; Franchi, M.; D’Adamo, R. Quality aspects of Crassostrea gigas (Thunberg, 1793) reared in the Varano Lagoon (southern Italy) in relation to marketability. J. Mar. Biol. Assoc. 2018, 98, 71–79. [Google Scholar] [CrossRef]
- Jensen, A.C.; Collins, K.; Lockwood, A.P. Artificial Reefs in European Seas. Available online: https://www.springer.com/gp/book/9780792358459 (accessed on 16 March 2021).
- MathWorks Inc., MATLAB (R2020b). Available online: https://www.mathworks.com/products/matlab.html?s_tid=srchtitle (accessed on 16 March 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2020; ISBN 3900051003. [Google Scholar]
- Quantum GIS Development Team. Quantum GIS Geographic Information System; QGIS: New York, NY, USA, 2020. [Google Scholar]
- Prioli, G. Censimento Nazionale Sulla Molluschicoltura Del Consorzio Unimar. Available online: http://www.unimar.it/wp-content/uploads/2017/04/18.-volume-Molluschicoltura.pdf (accessed on 16 March 2021).
- Salgado-Hernanz, P.M.; Racault, M.-F.; Font-Muñoz, J.S.; Basterretxea, G. Trends in phytoplankton phenology in the Mediterranean Sea based on ocean-colour remote sensing. Remote Sens. Environ. 2019, 221, 50–64. [Google Scholar] [CrossRef]
- Grangeré, K.; Ménesguen, A.; Lefebvre, S.; Bacher, C.; Pouvreau, S. Modelling the influence of environmental factors on the physiological status of the Pacific oyster Crassostrea gigas in an estuarine embayment; The Baie des Veys (France). J. Sea Res. 2009, 62, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Bernardi Aubry, F.; Acri, F. Phytoplankton seasonality and exchange at the inlets of the Lagoon of Venice (July 2001–June 2002). J. Mar. Syst. 2004, 51, 65–76. [Google Scholar] [CrossRef]
- Dupuy, C.; Vaquer, A.; Lam-Höai, T.; Rougier, C.; Mazouni, N.; Lautier, J.; Collos, Y.; Le Gall, S. Feeding rate of the oyster Crassostrea gigas in a natural planktonic community of the Mediterranean Thau Lagoon. Mar. Ecol. Prog. Ser. 2000, 205, 171–184. [Google Scholar] [CrossRef]
- Caroppo, C.; Fiocca, A.; Sammarco, P.; Magazzu, G. Seasonal variations of nutrients and phytoplankton in the coastal SW Adriatic Sea (1995–1997). Bot. Mar. 1999, 42, 389–400. [Google Scholar] [CrossRef]
- Volpe, G.; Nardelli, B.B.; Cipollini, P.; Santoleri, R.; Robinson, I.S. Seasonal to interannual phytoplankton response to physical processes in the Mediterranean Sea from satellite observations. Remote Sens. Environ. 2012, 117, 223–235. [Google Scholar] [CrossRef]
- O’Reilly, J.E.; Maritorena, S.; Mitchell, B.G.; Siegel, D.A.; Carder, K.L.; Garver, S.A.; Kahru, M.; McClain, C. Ocean color chlorophyll algorithms for SeaWiFS. J. Geophys. Res. Ocean. 1998, 103, 24937–24953. [Google Scholar] [CrossRef] [Green Version]
- Gregg, W.W.; Casey, N.W. Global and regional evaluation of the SeaWiFS chlorophyll data set. Remote Sens. Environ. 2004, 93, 463–479. [Google Scholar] [CrossRef]
- Zheng, G.; DiGiacomo, P.M. Remote sensing of chlorophyll-a in coastal waters based on the light absorption coefficient of phytoplankton. Remote Sens. Environ. 2017, 201, 331–341. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Han, E.; Wilberg, M.J.; Lee, W.C.; Choi, K.-S.; Kang, C.-K. Physiological processes and gross energy budget of the submerged longline-cultured Pacific oyster Crassostrea gigas in a temperate bay of Korea. PLoS ONE 2018, 13, e0199752. [Google Scholar] [CrossRef]
- Roncarati, A.; Felici, A.; Dees, A.; Leila, F.; Paolo, M. Trials on Pacific oyster (Crassostrea gigas Thunberg) rearing in the middle Adriatic Sea by means of different trays. Aquac. Int. 2010, 18, 35–43. [Google Scholar] [CrossRef]
- Cozzi, S.; Giani, M. River water and nutrient discharges in the Northern Adriatic Sea: Current importance and long term changes. Cont. Shelf Res. 2011, 31, 1881–1893. [Google Scholar] [CrossRef]
- Suquet, M.; Malo, F.; Quere, C.; Ledu, C.; Le Grand, J.; Benabdelmouna, A. Gamete quality in triploid Pacific oyster (Crassostrea gigas). Aquaculture 2016, 451, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Duchemin, M.B.; Fournier, M.; Auffret, M. Seasonal variations of immune parameters in diploid and triploid Pacific oysters, Crassostrea gigas (Thunberg). Aquaculture 2007, 264, 73–81. [Google Scholar] [CrossRef]
- Hawkins, A.J.S.; Magoulas, A.; Héral, M.; Bougrier, S.; Naciri-Graven, Y.; Day, A.J.; Kotoulas, G. Separate effects of triploidy, parentage and genomic diversity upon feeding behaviour, metabolic efficiency and net energy balance in the pacific oyster Crassostrea gigas. Genet. Res. 2000, 76, 273–284. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Bank Aquaculture Zoning, Site Selection and Area Management under the Ecosystem Approach to Aquaculture. Available online: http://www.fao.org/3/i6992e/i6992e.pdf (accessed on 16 March 2021).
- European Union. Directive 2014/89/EU of the European Parliment and of the Council of 23 July 2014 Establishing a Framework for Maritime Spatial Planning; European Union: Bruxelles, Belgium, 2014. [Google Scholar]
- Lourguioui, H.; Brigolin, D.; Boulahdid, M.; Pastres, R. A perspective for reducing environmental impacts of mussel culture in Algeria. Int. J. Life Cycle Assess. 2017, 22, 1266–1277. [Google Scholar] [CrossRef]
- Lindahl, O.; Hart, R.; Hernroth, B.; Kollberg, S.; Loo, L.-O.; Olrog, L.; Rehnstam-Holm, A.-S.; Svensson, J.; Svensson, S.; Syversen, U. Improving Marine Water Quality by Mussel Farming: A Profitable Solution for Swedish Society. AMBIO J. Hum. Environ. 2005, 34, 131–138. [Google Scholar] [CrossRef]
- Guéguen, M.; Amiard, J.-C.; Arnich, N.; Badot, P.-M.; Claisse, D.; Guérin, T.; Vernoux, J.-P. Shellfish and Residual Chemical Contaminants: Hazards, Monitoring, and Health Risk Assessment Along French Coasts Marielle. Rev. Environ. Contam. Toxicol. 2011, 213, 61–88. [Google Scholar] [CrossRef]
- Galli, G.; Solidoro, C.; Lovato, T. Marine Heat Waves Hazard 3D Maps and the Risk for Low Motility Organisms in a Warming Mediterranean Sea. Front. Mar. Sci. 2017, 4, 136. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, K.; De Young, C.; Soto, D. Climate Change Implications for Fisheries and Aquaculture Overview of Current Scientific Knowledge; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; ISBN 9789251063477. [Google Scholar]
- Soto-Navarro, J.; Jordá, G.; Amores, A.; Cabos, W.; Somot, S.; Sevault, F.; Macías, D.; Djurdjevic, V.; Sannino, G.; Li, L.; et al. Evolution of Mediterranean Sea water properties under climate change scenarios in the Med-CORDEX ensemble. Clim. Dyn. 2020, 54, 2135–2165. [Google Scholar] [CrossRef] [Green Version]






| Parameter | Value | Reference |
|---|---|---|
| Arrhenius temperature TA | 5800 K | [35] |
| Rate of decrease at lower boundary (TAL) | 75,000 K | [35] |
| Rate of decrease at upper boundary (TAH) | 30,000 K | [35] |
| Critical lower limit (TL) | 276 K | [31] |
| Critical ingestion upper limit (TH) | 298 K | [31] |
| Critical respiration upper limit (TH1) | 305 K | [31] |
| Half-saturation coefficient (XK) | 12.6 | This study |
| Max. surface area-specific ingestion (JXm) | 560 J/cm2 d | [35] |
| Assimilation efficiency (ae) | 0.75 | [35] |
| Volume-specific maintenance costs (p_M) | 24 J cm3/d | [35] |
| Max. reserve density (Em) | 2295 J/cm3 | [35] |
| Cost for growth (Eg) | 1900 J/cm3 | [35] |
| Energy content of reserve (mu E) | 17,500 J/cm2 | [36] |
| Allocation fraction to somatic tissue (k) | 1 | * |
| Volume at puberty (Vp) | n/a | [35] |
| Reproduction efficiency (KR) | 0 | * |
| Gonadosomatic index (GSI) | 0 | * |
| Shape coefficient (δm) | 0.225 ± 0.03 | This study |
| Wet meat weight to dry meat weight converter | 0.2 | [37] |
| Dry meat weight to total weight equation | 23.8 Wmd + 6 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertolini, C.; Brigolin, D.; Porporato, E.M.D.; Hattab, J.; Pastres, R.; Tiscar, P.G. Testing a Model of Pacific Oysters’ (Crassostrea gigas) Growth in the Adriatic Sea: Implications for Aquaculture Spatial Planning. Sustainability 2021, 13, 3309. https://doi.org/10.3390/su13063309
Bertolini C, Brigolin D, Porporato EMD, Hattab J, Pastres R, Tiscar PG. Testing a Model of Pacific Oysters’ (Crassostrea gigas) Growth in the Adriatic Sea: Implications for Aquaculture Spatial Planning. Sustainability. 2021; 13(6):3309. https://doi.org/10.3390/su13063309
Chicago/Turabian StyleBertolini, Camilla, Daniele Brigolin, Erika M. D. Porporato, Jasmine Hattab, Roberto Pastres, and Pietro Giorgio Tiscar. 2021. "Testing a Model of Pacific Oysters’ (Crassostrea gigas) Growth in the Adriatic Sea: Implications for Aquaculture Spatial Planning" Sustainability 13, no. 6: 3309. https://doi.org/10.3390/su13063309