

Alien Macroalgal Rearrangement in the Soft Substrata of the Venice Lagoon (Italy): Impacts, Threats, Time and Future Trends
 , , , and
, , , and
Abstract
:1. Introduction
Allochthonous Species
2. Materials and Methods
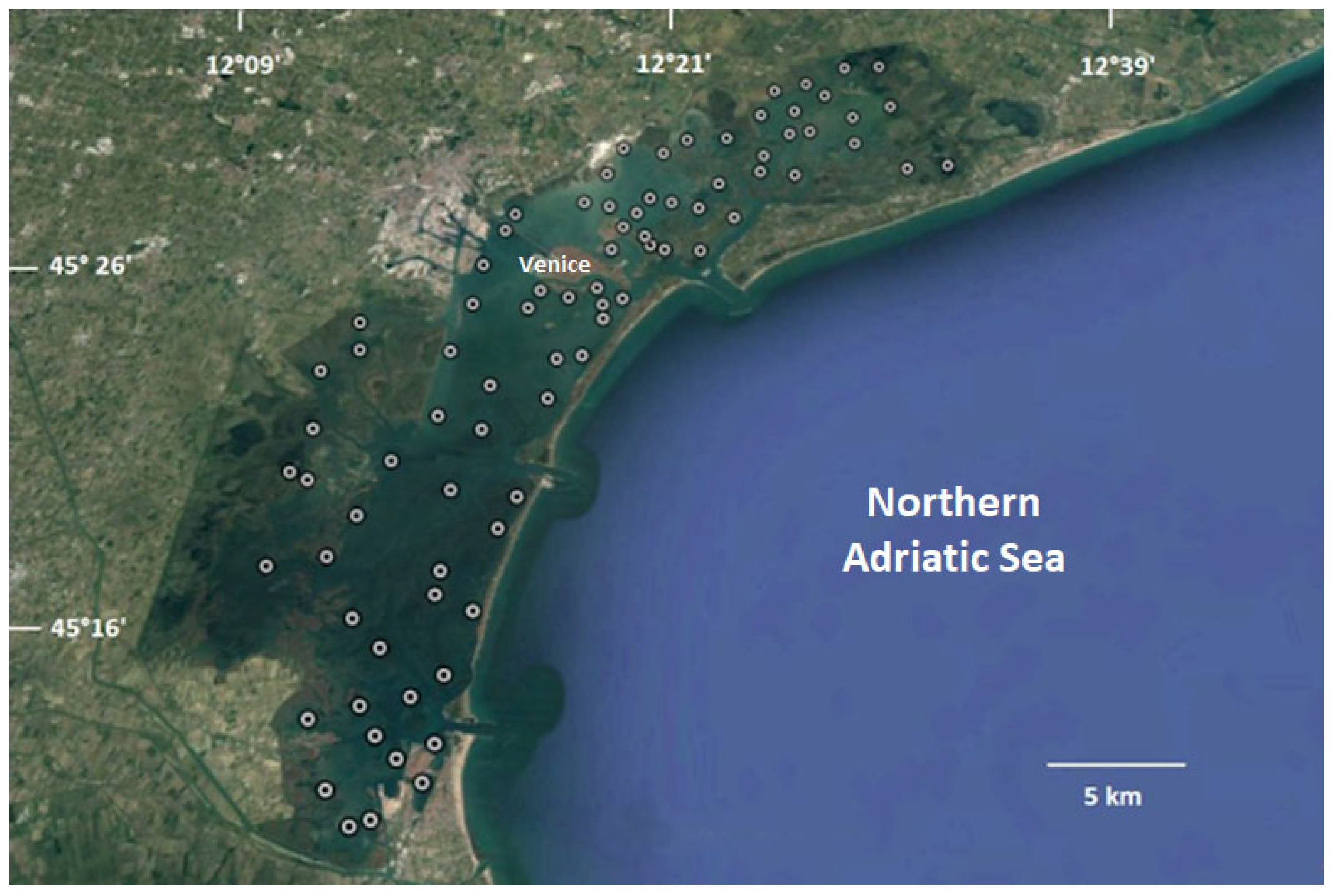
2.1. Study Area
2.2. Macrophyte Sampling
2.3. Molecular Analyses
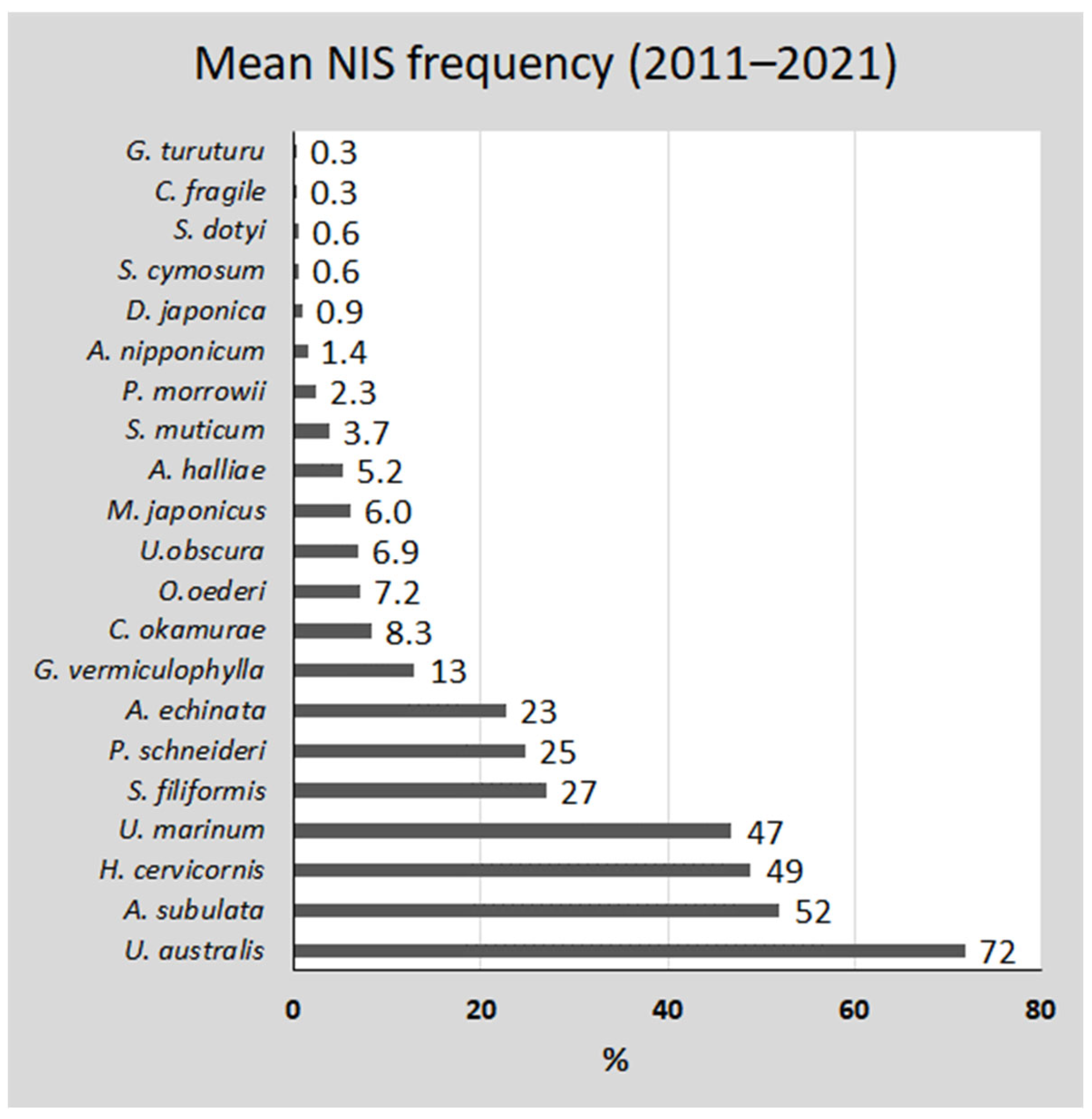
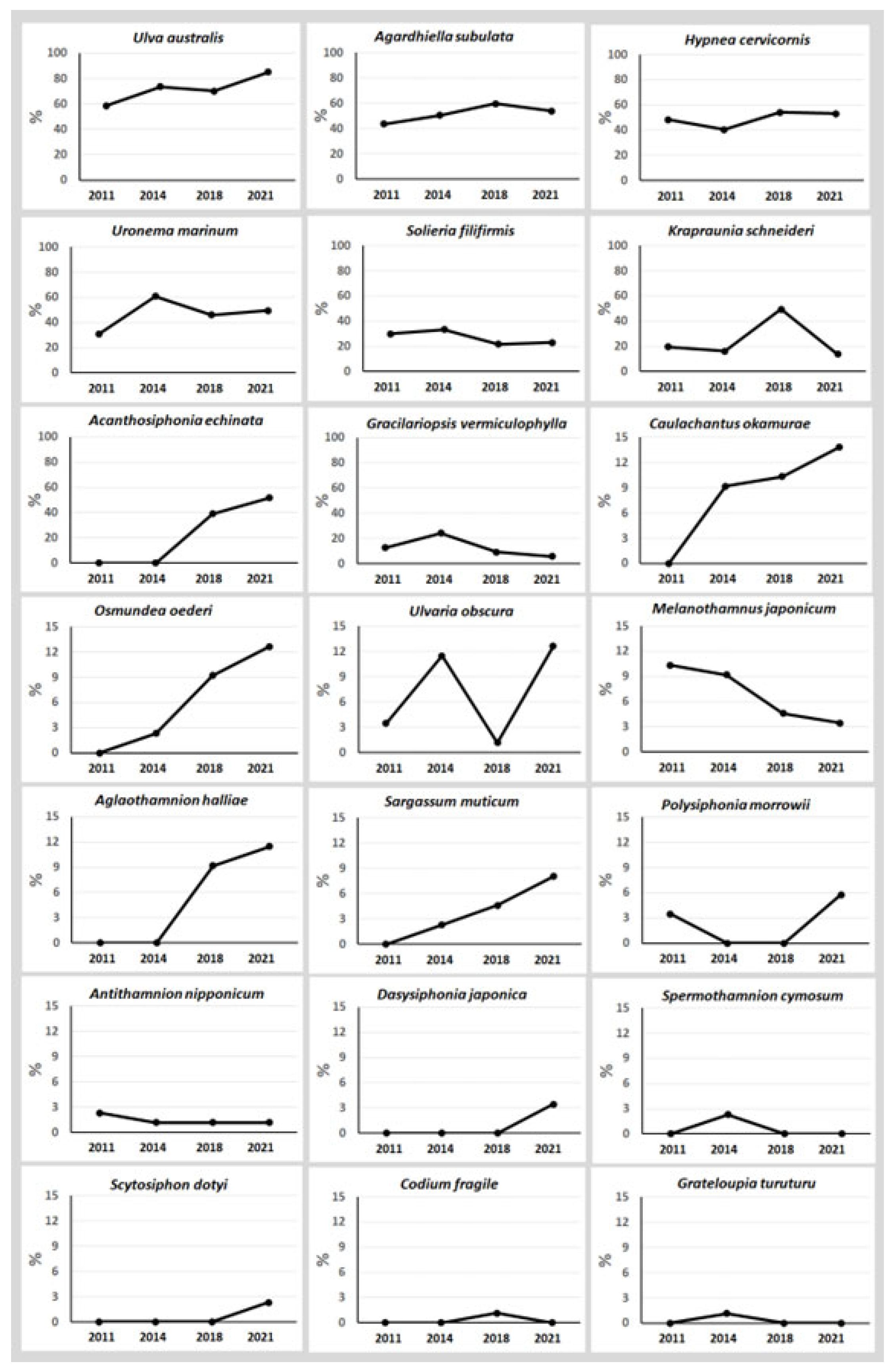
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Streftaris, N.; Zenetos, A.; Papathanassiou, E. Globalisation in marine ecosystems: The story of non-indigenous marine species across European seas. Oceanogr. Mar. Biol. 2005, 43, 419–453. [Google Scholar] [CrossRef]
- Savini, D.; Occhipinti–Ambrogi, A.; Marchini, A.; Tricarico, E.; Gherardi, F.; Olenin, S.; Gollasch, S. The top 27 animal alien species introduced into Europe for aquaculture and related activities. J. Appl. Ichthyol. 2010, 26, 1–7. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M.C.; Giaccone, G.; et al. Alien species along the Italian coasts: An overview. Biol. Invasions 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppakoski, E.; Çinar, M.E.; Ozturk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of marine invasive alien species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Tsiamis, K.I.; Azzurro, E.; Bariche, M.; Çinar, M.E.; Crocetta, F.; De Clerck, O.; Galil, B.; Gómez, F.; Hoffman, R.; Jensen, K.R.; et al. Prioritizing marine invasive alien species in the European Union through horizon scanning. Aquat. Conserv. 2020, 30, 794–845. [Google Scholar] [CrossRef]
- Tsirintanis, K.; Azzurro, E.; Crocetta, F.; Dimiza, M.; Froglia, C.; Gerovasileiou, V.; Langeneck, J.; Mancinelli, G.; Rosso, A.; Stern, N.; et al. Bioinvasion impacts on biodiversity, ecosystem services, and human health in the Mediterranean Sea. Aquat. Invasions 2022, 17, 308–352. [Google Scholar] [CrossRef]
- Çinar, M.E.; Bilecenoğlu, M.; Yokeş, M.B.; Öztürk, B.; Taşkin, E.; Bakir, K.; Doğan, A.; Açik, Ş. Current status (as of end of 2020) of marine alien species in Turkey. PLoS ONE 2021, 16, e0251086. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A.S. Building the European Alien Species Information Network (EASIN): A novel approach for the exploration of distributed alien species data. BioInvasions Rec. 2012, 1, 235–245. [Google Scholar] [CrossRef]
- Sousa, R.; Novais, A.; Costa, R.; Strayer, D.L. Invasive bivalves in fresh waters: Impacts from individuals to ecosystems and possible control strategies. Hydrobiologia 2014, 735, 233–251. [Google Scholar] [CrossRef]
- Chiesa, S.; Lucentini, L.; Freitas, R.; Marzano, F.N.; Breda, S.; Figueira, E.; Caill-Milly, N.; Herbert, R.; Soares, A.M.V.M.; Argese, E. A history of invasion: COI phylogeny of Manila clam Ruditapes philippinarum in Europe. Fish. Res. 2017, 186, 25–35. [Google Scholar] [CrossRef]
- Pranovi, F.; Franceschini, G.; Casale, M.; Zucchetta, M.; Giovanardi, O.; Torricelli, P. An ecological imbalance induced by a non-native species: The Manila clam in the Venice Lagoon. Biol. Invasions 2006, 8, 595–609. [Google Scholar] [CrossRef]
- Kim, S.L.; Lee, H.G.; Kang, S.M.; Yu, O.H. The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem. Sustainability 2020, 12, 4205. [Google Scholar] [CrossRef]
- Carol, J.; Benejam, L.; Benito, J.; García-Berthou, E. Growth and diet of European catfish (Silurus glanis) in early and late invasion stages. Fundam. Appl. Limnol. 2009, 74, 317–328. [Google Scholar] [CrossRef]
- Syvaranta, J.; Cucherousset, J.; Kopp, D.; Crivelli, A.; Cereghino, R.; Santoul, F. Dietary breadth and trophic position of introduced European catfish (Silurus glanis) in the river Tarn (Garonne river basin), southwest France. Aquat. Biol. 2010, 8, 137–144. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, R.; Cucherousset, J.; García-Berthou, E.; Kirk, R.; Peeler, E.; Stakénas, S. Voracious invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish Fish. 2009, 10, 252–282. [Google Scholar] [CrossRef]
- Fuentes, V.L.; Angel, D.L.; Bayha, K.M.; Atienza, D.; Edelist, D.; Bordehore, C.; Gili, J.-M.; Purcell, J.E. Blooms of the invasive ctenophore, Mnemiopsis leidyi, span the Mediterranean Sea in 2009. Hydrobiologia 2010, 645, 23–37. [Google Scholar] [CrossRef]
- Mancinelli, G.; Chainho, P.; Cilenti, L.; Falco, S.; Kapiris, K.; Katselis, G.; Ribeiro, F. The Atlantic blue crab Callinectes sapidus in southern European coastal waters: Distribution, impact and prospective invasion management strategies. Mar. Pollut. Bull. 2017, 119, 5–11. [Google Scholar] [CrossRef]
- Scalici, M.; Chiesa, S.; Mancinelli, G.; Rontani, P.M.; Voccia, A.; Nonnis Marzano, F. Euryhaline aliens invading Italian inland waters: The Case of the Atlantic Blue Crab Callinectes sapidus Rathbun, 1896. Appl. Sci. 2022, 12, 4666. [Google Scholar] [CrossRef]
- Verlaque, M. Check-list of the macroalgae of Thau lagoon (Hérault, France): A hot spot of marine species introduction in Europe. Oceanol. Acta 2001, 24, 29–49. [Google Scholar] [CrossRef]
- Verlaque, M.; Ruitton, S.; Mineur, F.; Boudouresque, C.F. Macrophytes. In CIESM Atlas of Exotic Species in the Mediterranean; CIESM Publishers: Paris, France, 2015; Volume 4, p. 364. [Google Scholar]
- Boudouresque, C.; Klein, J.C.; Ruitton, S.; Verlaque, M. Biological invasion: The Thau Lagoon, a Japanese Biological Island in the Mediterranean Sea. In Global Change: Mankind-Marine Environment Interactions; Ceccaldi, H.J., Dekeyser, I., Girault, M., Stora, G., Eds.; Springer Science: Dordrecht, The Netherlands, 2011; pp. 151–156. [Google Scholar]
- Marchini, A.; Ferrario, J.; Sfriso, A.; Occhipinti-Ambrogi, A. Current status and trends of biological invasions in the Lagoon of Venice, a hotspot of marine NIS introductions in the Mediterranean Sea. Biol. Invasions 2015, 17, 2943–2962. [Google Scholar] [CrossRef]
- Camatti, E.; Pansera, M.; Bergamasco, A. The Copepod Acartia tonsa Dana in a microtidal mediterranean lagoon: History of a successful invasion. Water 2019, 11, 1200. [Google Scholar] [CrossRef]
- Petrocelli, A.; Cecere, E.; Verlaque, M. Alien marine macrophytes in transitional water systems: New entries and reappearances in a Mediterranean coastal basin. Bioinvasions Rec. 2013, 2, 177–184. [Google Scholar] [CrossRef]
- Giangrande, A.; Arduini, D.; Borghese, J.; Del Pasqua, M.; Lezzi, M.; Petrocelli, A.; Cecere, E.; Longo, C.; Pierri, C. Macrobenthic success of non-indigenous species related to substrate features in the Mar Grande of Taranto, Italy (Mediterranean Sea). Bioinvasions Rec. 2021, 10, 238–256. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A. Biotic invasions in a Mediterranean Lagoon. Biol. Invasions 2000, 2, 165–176. [Google Scholar] [CrossRef]
- Available online: www.teknoring.com/news/trasporti-e-automotive/stop-grandi-navi-venezia-concorso-idee (accessed on 20 February 2023).
- Pessa, G.; Fischetti, D.; Saggese, A. Piano D’uso delle Aree in Concessione per Venericoltura. Aggiornamento. [Use Plan of the Areas under Concession for Clam Farming. Update]; San Servolo S.r.l. Servizi Metropolitani di Venezia; Gestione Risorse Lagunari Venezia: Venice, Italy, 2018; pp. 1–99. [Google Scholar]
- Turolla, E.; Boffo, L.; Rossetti, E. Venericoltura, un’eccellenza nazionale ed europea a rischio per la carenza di seme [Venericulture, a national and European excellence at risk due to seed shortage]. Il Pesce 2019, 19, 58–65. [Google Scholar]
- Sfriso, A.; Buosi, A.; Wolf, M.A.; Sfriso, A.A. Invasion of alien macroalgae in the Venice Lagoon, a pest or a resource? Aquat. Invasions 2020, 15, 245–270. [Google Scholar] [CrossRef]
- Wolf, M.A.; Buosi, A.; Sfriso, A. First record of Acanthosiphonia echinata (Rhodomelaceae, Rhodophyta) in the Mediterranean Sea, molecular and morphological characterization. Bot. Mar. 2020, 63, 241–245. [Google Scholar] [CrossRef]
- Petrocelli, A.; Wolf, A.M.; Cecere, E.; Sciuto, K.; Sfriso, A. Settlement and spreading of the introduced seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea. Diversity 2020, 12, 129. [Google Scholar] [CrossRef]
- Glasby, T.M.; Connell, S.D.; Holloway, M.G.; Hewitt, C.L. Nonindigenous biota on artificial structures: Could habitat creation facilitate biological invasions? Mar. Biol. 2007, 151, 887–895. [Google Scholar] [CrossRef]
- Marchini, A.; Sconfietti, R.; Krapp-Schickel, T. Role of the artificial structures on biodiversity: The case of arthropod fauna in the N-Adriatic lagoons. Studi Trent. Sci. Nat. Acta Biol. 2007, 83, 27–31. [Google Scholar]
- Airoldi, L.; Turon, Z.; Perkol-Finkel, S.; Rius, M. Corridors for aliens but not for natives: Effects of marine urban sprawl at a regional scale. Divers. Distrib. 2015, 21, 755–768. [Google Scholar] [CrossRef]
- Miozzi, E. Venezia nei Secoli [Venice through the Ages]; Officine Grafiche Trevisan: Castelfranco Veneto, Italy, 1969; Volume 3. [Google Scholar]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Marcomini, A. Recording the occurrence of trophic level changes in the lagoon of Venice over the ‘90s. Environ. Int. 2005, 31, 993–1001. [Google Scholar] [CrossRef]
- Melaku Kanu, D.; Campostrini, P.; Dalla Riva, S.; Pastres, R.; Pizzo, L.; Rossetto, L.; Solidoro, C. Addressing sustainability of clam farming in the Venice Lagoon. Ecol. Soc. 2011, 16, 26. [Google Scholar] [CrossRef]
- Cucco, A.; Umgiesser, G. Modelling the Venice Lagoon Residence Time. Ecol. Model. 2006, 193, 34–51. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Facca, C.; Sfriso, A.A. Role of environmental factors in affecting macrophyte dominance in transitional environments: The Italian Lagoons as a study case. Mar. Ecol. 2017, 38, e12414. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Bonometto, A.; Boscolo, R. Compliance of the Macrophyte Quality index (MaQI) with the WFD (2000/60/EC) and ecological status assessment in transitional areas: The Venice lagoon as study case. Ecol. Indic. 2014, 46, 536–547. [Google Scholar] [CrossRef]
- Guiry and Guiry. 2023. Available online: www.algaebase.org (accessed on 27 March 2023).
- Phillips, J.A. Field, anatomical and developmental studies on southern Australian species of Ulva (Ulvaceae, Chlorophyta). Aust. Syst. Bot. 1988, 1, 411–456. [Google Scholar] [CrossRef]
- Furnari, G.; Cormaci, M.; Serio, D. Catalogue of the benthic marine macroalgae of the Italian coast of the Adriatic Sea. Bocconea 1999, 12, 1–214. [Google Scholar]
- Furnari, G.; Giaccone, G.; Cormaci, M.; Alongi, G.; Serio, D. Biodiversità marina delle coste italiane: Catalogo del macrofitobenthos [Marine biodiversity of the Italian coasts: Catalog of macrophytobenthos]. Biol. Mar. Med. 2003, 10, 1–484. [Google Scholar]
- Sfriso, A. Coexistence of Ulva rigida and Ulva laetevirens (Ulvales, Chlorophyta) in Venice Lagoon and other Italian transitional and marine environments. Bot. Mar. 2010, 53, 9–18. [Google Scholar] [CrossRef]
- Manghisi, A.; Armeli Minicante, S.; Bertuccio, C.; Morabito, M.; Torricelli, P.; Genovese, G. A cryptic alien seaweed spreading in Mediterranean coastal lagoons. Transit. Water Bull. 2011, 5, 1–7. [Google Scholar] [CrossRef]
- Wolf, M.A.; Sciuto, K.; Andreoli, C.; Moro, I. Ulva (Chlorophyta, Ulvales) biodiversity in the North Adriatic Sea (Mediterranean, Italy): Cryptic species and new introductions. J. Phycol. 2012, 48, 1510–1521. [Google Scholar] [CrossRef]
- Coucerio, L.; Cremades, J.; Barreiro, R. Evidence for multiple introductions of the Pacific green alga Ulva australis Areschoug (Ulvales, Chlorophyta) to the Iberian Peninsula. Bot. Mar. 2011, 54, 391–402. [Google Scholar] [CrossRef]
- Kraft, L.G.T.; Kraft, G.T.; Waller, R.F. Investigation into southern Australian Ulva (Ulvophyceae, Chlorophyta) taxonomy and molecular phylogeny indicate both cosmopolitan and endemic cryptic species. J. Phycol. 2010, 46, 1257–1277. [Google Scholar] [CrossRef]
- Womersley, H.B.S. The Marine Benthic Flora of Southern Australia. Part I. Adelaide: South Australian Government Printing Division; Taylor & Francis: Abingdon, UK, 1984; pp. 1–328. [Google Scholar]
- Sfriso, A.; Curiel, D.; Rismondo, A. The Venice Lagoon. In Flora and Vegetation of the Italian Transitional Water Systems; Cecere, E., Petrocelli, A., Izzo, G., Sfriso, A., Eds.; CoRiLa Multigraf Spinea: Venice, Italy, 2009; pp. 17–80. [Google Scholar]
- Thomsen, M.S.; Gurgel, C.F.D.; Fredericq, S.; McGlathery, K.J. Gracilaria vermiculophylla (Rhodophyta, Gracilariales) in Hog Island Bay, Virginia: A cryptic alien and invasive macroalga and taxonomic correction. J. Phycol. 2005, 42, 139–141. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Staehr, P.A.; Nyberg, C.D.; Schwaerter, S.; Krause-Jensen, D.; Silliman, B.R. Gracilaria vermiculophylla (Ohmi) Papenfuss, 1967 (Rhodophyta, Gracilariaceae) in northern Europe, with emphasis on Danish conditions, and what to expect in the future. Aquat. Invasions 2007, 2, 83–94. [Google Scholar] [CrossRef]
- Krueger-Hadfield, S.A.; Magill, C.L.; Bunker, F.S.P.D.; Mieszkowska, N.; Sotka, E.E.; Maggs, C.A. When Invaders Go Unnoticed: The Case of Gracilaria vermiculophylla in the British Isles. Cryptogam. Algol. 2017, 38, 379–400. [Google Scholar] [CrossRef]
- Curiel, D.; Bellemo, G.; Scattolin, M.; La Rocca, B.; Marzocchi, M. Ritrovamento in laguna di Venezia di specie algali nuove per l’Adriatico e rare per il Mediterraneo [Discovery in the Venice Lagoon of algal species new to the Adriatic Sea and rare for the Mediterranean Sea]. Inf. Bot. Ital. 2003, 35, 7–11. [Google Scholar]
- Curiel, D.; Bellemo, G.; Checchin, E.; Dri, C.; Miotti, C.; Marzocchi, M. Segnalazione di nuove macroalghe per la laguna di Venezia [Reporting of new macroalgae for the Venice Lagoon]. Lavori Soc. Ven. Sci. Nat. 2005, 30, 41–44. [Google Scholar]
- Curiel, D.; Bellemo, G.; La Rocca, B.; Scattolin, M.; Marzocchi, M. Note su specie algali nuove per la laguna di Venezia [Notes on algal species new to the Venice Lagoon]. Lavori Soc. Ven. Sci. Nat. 2001, 26, 101–102. [Google Scholar]
- Bellemo, G.; Curiel, D.; Marzocchi, M.; Iuri, M.; Pavan, M.; Vecchiato, N.; Scattolin, M. Aggiornamento di nuove specie algali per la laguna di Venezia [Update of new algal species for the Venice Lagoon]. Lavori Soc. Ven. Sci. Nat. 1999, 24, 55–66. [Google Scholar]
- Curiel, D.; Bellemo, G.; Marzocchi, M. New records of marine algae in the Lagoon of Venice. Plant Biosyst. 1996, 130, 352. [Google Scholar] [CrossRef]
- Athanasiadis, A. Phycologia Europaea Rhodophyta; Published and Distributed by the Author: Thessaloniki, Greece, 2016; Volume 2, pp. 763–1504. [Google Scholar]
- Kim, H.-S.; Lee, I.K. Rhodophyta: Florideophyceae: Ceramiales: Ceramiaceae I (Non-corticate species). In Algal Flora of Korea; National Institute of Biological Resources: Incheon, Republic of Korea, 2012; Volume 4. [Google Scholar]
- Armeli-Minicante, S. Spermothamnion cymosum (Harvey) De Toni. Prima segnalazione della specie in Adriatico [First report of the species in the Adriatic]. Boll. Mus. Storia Nat. Venezia 2013, 64, 143–144. [Google Scholar]
- Gargiulo, M.G.; De Masi, F.; Tripodi, G. Sargassum muticum (Yendo) Fensholt (Phaeophyta, Fucales) is spreading in the Lagoon of Venice (Northern Adriatic Sea). Giorn. Bot. Ital. 1992, 126, 259. [Google Scholar]
- Tolomio, C. Prima segnalazione di Grateloupia doryphora (Mont.) Howe (Rhodophyceae) nella laguna di Venezia [First record of Grateloupia doryphora (Mont.) Howe (Rhodophyceae) in the Venice lagoon]. Lavori Soc. Ven. Sci. Nat. 1993, 18, 215–220. [Google Scholar]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A.; Gollasch, S.; Minchin, D.; Narščius, A.; Ojaveer, H.; Olenin, S. International arrivals: Widespread bioinvasions in European seas. Ethol. Ecol. Evol. 2014, 26, 152–171. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A. Biopollution by invasive marine non-indigenous species: A review of potential adverse ecological effects in a changing climate. Int. J. Environ. Res. Public Health 2021, 8, 4268. [Google Scholar] [CrossRef]
- Tamburini, M.; Keppel, E.; Marchini, A.; Repetto, M.F.; Ruiz, M.G.; Ferrario, J.; Occhipinti-Ambrogi, A. Monitoring non-indigenous species in port habitats: First application of a standardized North American protocol in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 700730. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Tricarico, E.; Vilizzi, L.; Bilge, G.; Ekmekçi, F.G.; Filiz, H.; Giannetto, D.; İlhan, A.; Killi, N.; Kirankaya, Ş.G.; et al. Risk of invasiveness of non-native aquatic species in the eastern Mediterranean region under current and projected climate conditions. Eur. Zool. J. 2021, 88, 1130–1143. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Sciuto, K.; Wolf, M.; Tomio, Y.; Juhmani, A.-S.; Sfriso, A.A. Effect of ecological recovery on macrophyte dominance and production in the Venice Lagoon. Front. Mar. Sci. 2022, 9, 882463. [Google Scholar] [CrossRef]
- Nyberg, C.D.; Thomsen, M.S.; Wallentinus, I. Flora and fauna associated with the introduced red alga Gracilaria vermiculophylla. Eur. J. Phycol. 2009, 44, 395–403. [Google Scholar] [CrossRef]
- Ramus, A.P.; Silliman, R.B.; Thomsen, M.S.; Long, Z.T. An invasive foundation species enhances multifunctionality in a coastal ecosystem. Proc. Natl. Acad. Sci. USA 2017, 114, 8580–8585. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.A.; Lipcius, R.N. Non-native red alga Gracilaria vermiculophylla compensates for seagrass loss as blue crab nursery habitat in the emerging Chesapeake Bay ecosystem. PLoS ONE 2022, 17, e0267880. [Google Scholar] [CrossRef] [PubMed]
- EC 2014; Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. L 317/35. European Union: Maastricht, The Netherlands, 2014.




{kind=link}
{kind=link}
{kind=link}
| Alien Species in Soft Substrata of the Venice Lagoon | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N° | Alien Species | First Record in Venice | Number of Stations | % of Stations | Standing Crop 2014 (Sfriso et al., 2020) [30] | ||||||||||
| Study | Probable date | 2011 | 2014 | 2018 | 2021 | Mean | 2011 | 2014 | 2018 | 2021 | Mean | ||||
| 1 | U. australis | 2011 | <2007 | 51 | 64 | 61 | 74 | 63 | 59 | 74 | 70 | 85 | 72 | unknown | |
| 2 | A. subulata | 2003 | 2003 | 38 | 44 | 52 | 47 | 45 | 44 | 51 | 60 | 54 | 52 | 36,714 | tonnes FWT |
| 3 | H. cervicornis | 2011 | 2009 | 42 | 35 | 47 | 46 | 43 | 48 | 40 | 54 | 53 | 49 | 28,305 | tonnes FWT |
| 4 | U. marinum | 2014 | 2008 | 27 | 53 | 40 | 43 | 41 | 31 | 61 | 46 | 49 | 47 | 8.1 | tonnes FWT |
| 5 | S. filiformis | 2005 | 2003 | 26 | 29 | 19 | 20 | 24 | 30 | 33 | 22 | 23 | 27 | 3768 | tonnes FWT |
| 6 | K. schneideri | 2018 | <2007 | 17 | 14 | 43 | 12 | 22 | 20 | 16 | 49 | 14 | 25 | 398 | tonnes FWT |
| 7 | A. echinata | 2019 | 2014 | 0 | 0 | 34 | 45 | 20 | 0 | 0 | 39 | 52 | 23 | 0 | |
| 8 | G. vermiculophylla | 2008 | 2008 | 11 | 21 | 8 | 5 | 11 | 13 | 24 | 9.2 | 5.7 | 13 | 66,383 | tonnes FWT |
| 9 | C. okamurae | 2020 | 2014 | 0 | 8 | 9 | 12 | 7 | 0.0 | 9.2 | 10 | 13.8 | 8.3 | 0 | |
| 10 | U.obscura | 2002 | 2000 | 3 | 10 | 1 | 11 | 6 | 3.4 | 11.5 | 1.1 | 12.6 | 7.2 | 323 | tonnes FWT |
| 11 | M. japonicus | 2018 | 1998 | 9 | 8 | 4 | 3 | 6 | 10 | 9.2 | 4.6 | 3.4 | 6.9 | 272 | tonnes FWT |
| 12 | O. oederi | 2021 | 2014 | 0 | 2 | 8 | 11 | 5 | 0 | 2.3 | 9.2 | 12.6 | 6.0 | 0 | |
| 13 | A. halliae | 2018 | 2016 | 0 | 0 | 8 | 10 | 5 | 0 | 0 | 9.2 | 11.5 | 5.2 | <0.5 | kg FWT |
| 14 | S. muticum | 1992 | 1992 | 0 | 2 | 4 | 7 | 3 | 0 | 2.3 | 4.6 | 8.0 | 3.7 | 4825 | tonnes FWT |
| 15 | P. morrowii | 2001 | 1999 | 3 | 0 | 0 | 5 | 2 | 3.4 | 0 | 0.0 | 5.7 | 2.3 | 517 | tonnes FWT |
| 16 | A. nipponicum | 1996 | 1994 | 2 | 1 | 1 | 1 | 1 | 2.3 | 1.1 | 1.1 | 1.1 | 1.4 | 3.1 | tonnes FWT |
| 17 | D. japonica | 2005 | 1999 | 0 | 0 | 0 | 3 | 0.8 | 0 | 0 | 0 | 3.4 | 0.9 | + | |
| 18 | S. cymosum | 2013 | 2010 | 0 | 2 | 0 | 0 | 0.5 | 0 | 2.3 | 0 | 0 | 0.6 | + | |
| 19 | S. dotyi | 1996 | 1996 | 0 | 0 | 0 | 2 | 0.5 | 0 | 0 | 0 | 2.3 | 0.6 | 4775 | tonnes FWT |
| 20 | C. fragile | 1987 | 1978 | 0 | 0 | 1 | 0 | 0.3 | 0 | 0 | 1.1 | 0 | 0.3 | 1.25 | tonnes FWT |
| 21 | G. turuturu | 1993 | 1989 | 0 | 1 | 0 | 0 | 0.3 | 0 | 1.1 | 0 | 0 | 0.3 | 87 | tonnes FWT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sfriso, A.; Wolf, M.A.; Buosi, A.; Sciuto, K.; Sfriso, A.A. Alien Macroalgal Rearrangement in the Soft Substrata of the Venice Lagoon (Italy): Impacts, Threats, Time and Future Trends. Sustainability 2023, 15, 8256. https://doi.org/10.3390/su15108256
Sfriso A, Wolf MA, Buosi A, Sciuto K, Sfriso AA. Alien Macroalgal Rearrangement in the Soft Substrata of the Venice Lagoon (Italy): Impacts, Threats, Time and Future Trends. Sustainability. 2023; 15(10):8256. https://doi.org/10.3390/su15108256
Chicago/Turabian StyleSfriso, Adriano, Marion Adelheid Wolf, Alessandro Buosi, Katia Sciuto, and Andrea Augusto Sfriso. 2023. "Alien Macroalgal Rearrangement in the Soft Substrata of the Venice Lagoon (Italy): Impacts, Threats, Time and Future Trends" Sustainability 15, no. 10: 8256. https://doi.org/10.3390/su15108256