
Inhibition of Larval Development of Marine Copepods Acartia tonsa by Neonicotinoids

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acartia Tonsa Culturing
2.2. Chemicals
2.3. Toxicity Testing
2.4. Endpoints Measured
2.5. Chemical Analysis
2.6. Data Analysis
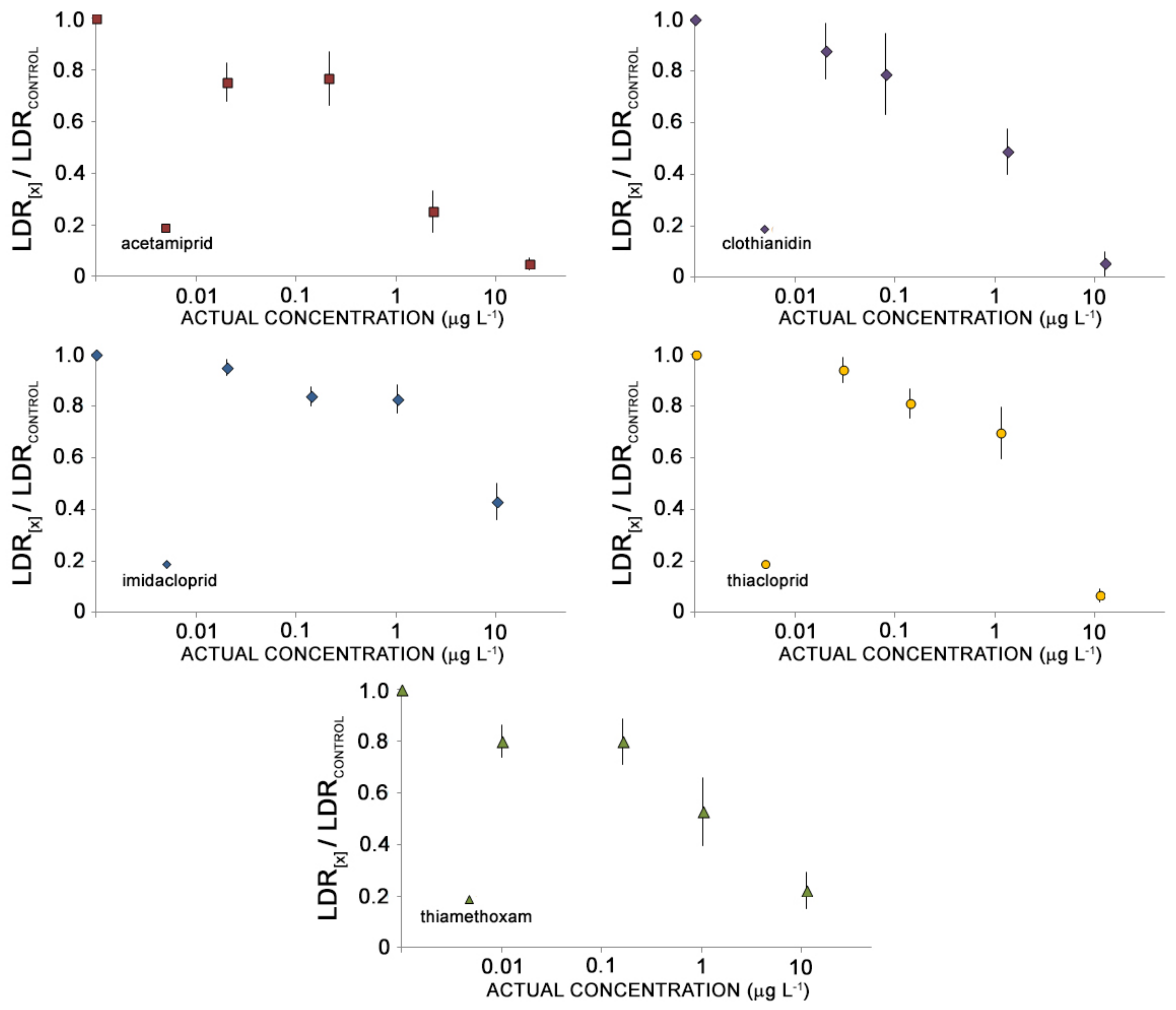
3. Results
3.1. Quality Assurance/Quality Control (QA/QC)
3.2. Toxicity Testing
4. Discussion
4.1. The Sensitivity of A. tonsa towards NEOs
4.2. Comparative Toxicity of NEOs
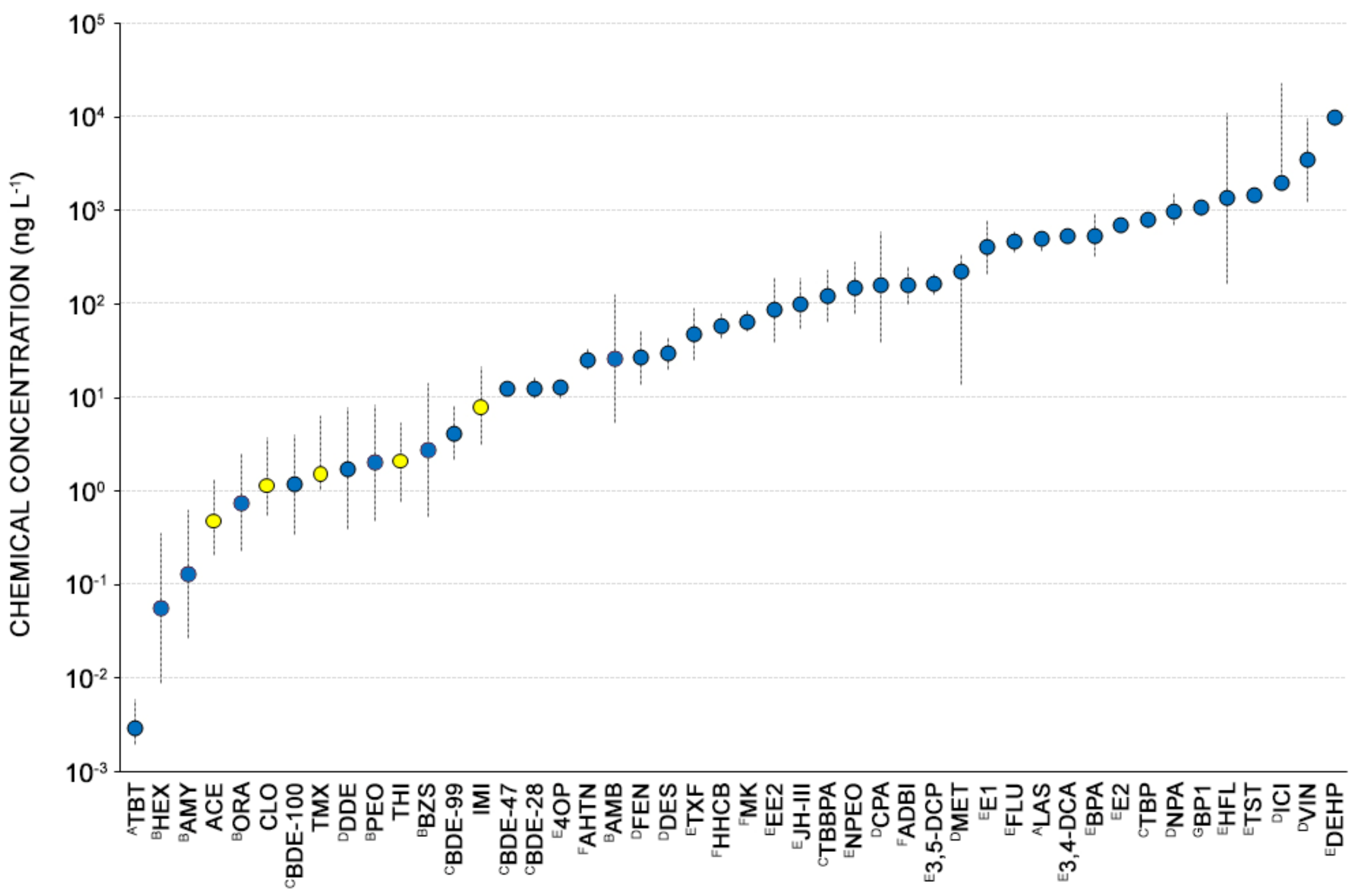
4.3. Environmental Significance of NEOs Toxicity toward Copepods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Belzunces, L.P.; Tchamitchian, S.; Brunet, J.L. Neural effects of insecticides in the honey bee. Apidologie 2012, 43, 348–370. [Google Scholar] [CrossRef] [Green Version]
- Blacquière, T.; Smagghe, G.; Van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (Neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Hladik, M.L.; Kolpin, D.W. First national-scale reconnaissance of neonicotinoid insecticides in streams across the USA. Environ. Chem. 2016, 13, 12. [Google Scholar] [CrossRef]
- Struger, J.; Grabuski, J.; Cagampan, S.; Sverko, E.; McGoldrick, D.; Marvin, C.H. Factors influencing the occurrence and distribution of neonicotinoid insecticides in surface waters of southern Ontario, Canada. Chemosphere 2017, 169, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Finnegan, M.C.; Baxter, L.R.; Maul, J.D.; Hanson, M.L.; Hoekstra, P.F. Comprehensive characterization of the acute and chronic toxicity of the neonicotinoid insecticide thiamethoxam to a suite of aquatic primary producers, invertebrates, and fish. Environ. Toxicol. Chem. 2017, 36, 2838–2848. [Google Scholar] [CrossRef] [Green Version]
- Raby, M.; Nowierski, M.; Perlov, D.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Acute toxicity of 6 neonicotinoid insecticides to freshwater invertebrates. Environ. Toxicol. Chem. 2018, 37, 1430–1445. [Google Scholar] [CrossRef]
- Van Wijngaarden, R.P.A.; Brock, T.C.M.; Van den Brink, P.J. Threshold levels for effects of insecticides in freshwater ecosystems: A review. Ecotoxicology 2005, 14, 355–380. [Google Scholar] [CrossRef] [PubMed]
- Gallen, C.; Paxman, C.; Thompson, K.; Mueller, J.; Da Silva, E.; Tracey, D.; Petus, C.; Devlin, M. Pesticide Monitoring in Inshore Waters of the Great Barrier Reef Using both Time-Integrated and Event Monitoring Techniques (2013–2014); The University of Queensland, The National Research Centre for Environmental Toxicology (Entox): Brisbane, Australia, 2014. [Google Scholar]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Bayo, F.; Goka, K.; Hayasaka, D. Contamination of the Aquatic Environment with Neonicotinoids and its Implication for Ecosystems. Front. Environ. Sci. 2016, 4, 71. [Google Scholar] [CrossRef]
- Hano, T.; Ito, K.; Ohkubo, N.; Sakaji, H.; Watanabe, A.; Takashima, K.; Sato, T.; Sugaya, T.; Matsuki, K.; Onduka, T.; et al. Occurrence of neonicotinoids and fipronil in estuaries and their potential risks to aquatic invertebrates. Environ. Pollut. 2019, 252, 205–215. [Google Scholar] [CrossRef]
- Osorio, D.; Bacon, J.P.; Whitington, P.M. The Evolution of Arthropod Nervous Systems: Modern insects and crustaceans diverged from an ancestor over 500 million years ago, but their neural circuitry retains many common features. Am. Sci. 1997, 85, 244–253. [Google Scholar]
- Smarandache-Wellmann, C.R. Arthropod neurons and nervous system. Curr. Biol. 2016, 26, R960–R965. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.; Conway, D.V.P.; Hunt, H.G. The role of copepods in the planktonic ecosystems of mixed and stratified waters of the European shelf seas. Hydrobiologia 1994, 292–293, 521–530. [Google Scholar] [CrossRef]
- Christoffersen, K.; Hansen, B.W.; Johansson, L.S.; Krog, E. Influence of LAS on marine calanoid copepod population dynamics and potential reproduction. Aquat. Toxicol. 2003, 63, 405–416. [Google Scholar] [CrossRef]
- Forget-Leray, J.; Landriau, I.; Minier, C.; Leboulenger, F. Impact of endocrine toxicants on survival, development, and reproduction of the estuarine copepod Eurytemora affinis (Poppe). Ecotoxicol. Environ. Saf. 2005, 60, 288–294. [Google Scholar] [CrossRef]
- Moeris, S.; Vanryckeghem, F.; Demeestere, K.; De Schamphelaere, K.A.C. Neonicotinoid insecticides from a marine perspective: Acute and chronic copepod testing and derivation of environmental quality standards. Environ. Toxicol. Chem. 2021, 40, 1353–1367. [Google Scholar] [CrossRef]
- Kusk, K.O.; Wollenberger, L. Towards an internationally harmonized test method for reproductive and developmental effects of endocrine disrupters in marine copepods. Ecotoxicology 2007, 16, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Wollenberger, L.; Breitholtz, M.; Ole Kusk, K.; Bengtsson, B.-E. Inhibition of larval development of the marine copepod Acartia tonsa by four synthetic musk substances. Sci. Total Environ. 2003, 305, 53–64. [Google Scholar] [CrossRef]
- Wollenberger, L.; Dinan, L.; Breitholtz, M. Brominated Flame Retardants: Activities in a crustacean development test and in an ecdysteroid screening assay. Environ. Toxicol. Chem. 2005, 24, 400. [Google Scholar] [CrossRef] [PubMed]
- Picone, M.; Distefano, G.G.; Marchetto, D.; Russo, M.; Vecchiato, M.; Gambaro, A.; Barbante, C.; Ghirardini, A.V. Fragrance materials (FMs) affect the larval development of the copepod Acartia tonsa: An emerging issue for marine ecosystems. Ecotoxicol. Environ. Saf. 2021, 215, 112146. [Google Scholar] [CrossRef]
- Picone, M.; Bergamin, M.; Delaney, E.; Volpi Ghirardini, A.; Kusk, K.O. Testing lagoonal sediments with early life stages of the copepod Acartia tonsa (Dana): An approach to assess sediment toxicity in the Venice Lagoon. Ecotoxicol. Environ. Saf. 2018, 147C, 217–227. [Google Scholar] [CrossRef]
- Thompson, D.A.; Lehmler, H.J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; Lefevre, G.H.; Peeples, T.L.; Poch, M.C.; Laduca, L.E.; et al. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nakamura, K.; Yokomizo, H. Relative robustness of NOEC and ECx against large uncertainties in data. PLoS ONE 2018, 13, e0206901. [Google Scholar] [CrossRef]
- Warne, M.S.J.; Van Dam, R. NOEC and LOEC data should no longer be generated or used. Australas. J. Ecotoxicol. 2008, 14, 1–5. [Google Scholar]
- ISO 16778:2015; Water Quality—Calanoid Copepod Early-Life Stage Test with Acartia tonsa. International Standard Organisation: Geneva, Switzerland, 2015; p. 37.
- Hook, S.E.; Doan, H.; Gonzago, D.; Musson, D.; Du, J.; Kookana, R.; Sellars, M.J.; Kumar, A. The impacts of modern-use pesticides on shrimp aquaculture: An assessment for north eastern Australia. Ecotoxicol. Environ. Saf. 2018, 148, 770–780. [Google Scholar] [CrossRef]
- Andersen, H.R.; Wollenberger, L.; Halling-Sørensen, B.; Kusk, K.O. Development of copepod nauplii to copepodites—A parameter for chronic toxicity including endocrine disruption. Environ. Toxicol. Chem. 2001, 20, 2821–2829. [Google Scholar] [CrossRef]
- Christensen, E.R.; Kusk, K.O.; Nyholm, N. Dose–response regressions for algal growth and similar continuous endpoints: Calculation of effective concentrations. Environ. Toxicol. Chem. 2009, 28, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Butcherine, P.; Kelaher, B.P.; Taylor, M.D.; Lawson, C.; Benkendorff, K. Acute toxicity, accumulation and sublethal effects of four neonicotinoids on juvenile Black Tiger Shrimp (Penaeus monodon). Chemosphere 2021, 275, 129918. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, J.S.; Darnell, K.M.; Blickley, T.M.; Romano, J.A.; Rittschof, D. Acute toxicity and sub-lethal effects of common pesticides in post-larval and juvenile blue crabs, Callinectes sapidus. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 5–14. [Google Scholar] [CrossRef]
- Song, M.Y.; Stark, J.D.; Brown, J.J. Comparative toxicity of four insecticides, including imidacloprid and tebufenozide, to four aquatic arthropods. Environ. Toxicol. Chem. 1997, 16, 2494–2500. [Google Scholar] [CrossRef]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Lethal and sublethal toxicity of neonicotinoid and butenolide insecticides to the mayfly, Hexagenia spp. Environ. Pollut. 2018, 238, 63–75. [Google Scholar] [CrossRef]
- Cavallaro, M.C.; Morrissey, C.A.; Headley, J.V.; Peru, K.M.; Liber, K. Comparative chronic toxicity of imidacloprid, clothianidin, and thiamethoxam to Chironomus dilutus and estimation of toxic equivalency factors. Environ. Toxicol. Chem. 2017, 36, 372–382. [Google Scholar] [CrossRef]
- Maloney, E.M.; Sykes, H.; Morrissey, C.; Peru, K.M.; Headley, J.V.; Liber, K. Comparing the Acute Toxicity of Imidacloprid with Alternative Systemic Insecticides in the Aquatic Insect Chironomus dilutus. Environ. Toxicol. Chem. 2020, 39, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Kusk, K.O.; Petersen, S. Acute and chronic toxicity of tributyltin and linear alkylbenzene sulfonate to the marine copepod Acartia tonsa. Environ. Toxicol. Chem. 1997, 16, 1629–1633. [Google Scholar] [CrossRef]
- Wollenberger, L. Toxicity Tests with Crustaceans for Detecting Sublethal Effects of Potential Endocrine Disrupting Chemicals; Technical University of Denmark: Kongens Lyngby, Denmark, 2005. [Google Scholar]
- Kusk, K.O.; Avdolli, M.; Wollenberger, L. Effect of 2,4-dihydroxybenzophenone (BP1) on early life-stage development of the marine copepod Acartia tonsa at different temperatures and salinities. Environ. Toxicol. Chem. 2011, 30, 959–966. [Google Scholar] [CrossRef]
- Andersen, H.R.; Halling-Sørensen, B.; Kusk, K.O. A parameter for detecting estrogenic exposure in the copepod Acartia tonsa. Ecotoxicol. Environ. Saf. 1999, 44, 56–61. [Google Scholar] [CrossRef]
- Maloney, E.M.; Taillebois, E.; Gilles, N.; Morrissey, C.A.; Liber, K.; Servent, D.; Thany, S.H. Binding properties to nicotinic acetylcholine receptors can explain differential toxicity of neonicotinoid insecticides in Chironomidae. Aquat. Toxicol. 2021, 230, 105701. [Google Scholar] [CrossRef] [PubMed]
- Smit, C.E.; Posthuma-Doodeman, C.J.A.M.; van Vlaardingen, P.L.A.; de Jong, F.M.W. Ecotoxicity of Imidacloprid to Aquatic Organisms: Derivation of Water Quality Standards for Peak and Long-Term Exposure. Hum. Ecol. Risk Assess. 2015, 21, 1608–1630. [Google Scholar] [CrossRef]
- Li, X.; Chen, J.; He, X.; Wang, Z.; Wu, D.; Zheng, X.; Zheng, L.; Wang, B. Simultaneous determination of neonicotinoids and fipronil and its metabolites in environmental water from coastal bay using disk-based solid-phase extraction and high-performance liquid chromatography–tandem mass spectrometry. Chemosphere 2019, 234, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.H.; Ng, K.T.; Lamphiere, A.; Cameron, T.C.; Bury, N.R.; Barron, L.P. Multicompartment and cross-species monitoring of contaminants of emerging concern in an estuarine habitat. Environ. Pollut. 2021, 270, 116300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CAS Number | Chemical Formula | Molar Mass (g mol−1) † | Water Solubility at 20 °C (mg L−1) † | Vapor Pressure at 20 °C (mPa) † | Log Kow † | Photolysis (t1/2 in d) † | Hydrolysis (t1/2 in d) † | |
|---|---|---|---|---|---|---|---|---|
| acetamiprid | 135410-20-7 | C10H11ClN4 | 222.7 | 2950 | 1.7 × 10−4 | 0.80 | 34 | stable |
| clothianidin | 210880-92-5 | C6H8ClN5O2S | 249.7 | 340 | 2.8 × 10−8 | 0.91 | <1 | stable |
| imidacloprid | 138261-41-3 | C9H10ClN5O2 | 255.7 | 610 | 4.0 × 10−7 | 0.57 | <1 | stable |
| thiacloprid | 111988-49-9 | C10H9ClN4S | 252.7 | 184 | 3.0 × 10−7 | 1.26 | stable | stable |
| thiamethoxam | 153719-23-4 | C8H10ClN5O3S | 291.7 | 4100 | 6.6 × 10−6 | −0.13 | 2.7 | stable |
| Effect-Concentrations for A. tonsa and 95% Confidence Interval | USEPA Aquatic Life Benchmarks | Environmental Quality Standards for Saltwater † | |||||
|---|---|---|---|---|---|---|---|
| EC10 | EC20 | EC50 | Acute | Chronic | AA-EQS | MAC-EQS | |
| acetamiprid | 0.05 (0.01–1.18) | 0.12 (0.01–1.75) | 0.73 (0.25–2.13) | 10.5 | 2.1 | - | - |
| clothianidin | 0.30 (0.04–2.11) | 0.56 (0.13–2.41) | 1.90 (0.99–3.63) | 11 | 0.05 | 0.05 | 0.23 |
| imidacloprid | 0.50 (0.11–2.22) | 1.33 (0.43–4.11) | 8.84 (5.13–15.24) | 0.385 | 0.01 | 0.002 | 0.065 |
| thiacloprid | 0.53 (0.21–1.34) | 0.88 (0.44–1.76) | 2.34 (1.49–3.69) | 18.9 | 0.97 | 0.0048 | 0.46 |
| thiamethoxam | 0.06 (0.01–0.71) | 0.18 (0.06–1.25) | 1.71 (0.61–4.80) | 17.5 | 0.74 | 0.016 | 5.2 |
| Species | Endpoint | Parameter | ACE | CLO | IMI | THI | TMX | Reference |
|---|---|---|---|---|---|---|---|---|
| Nitocra spinipes | Mortality | 96h-EC50 | - | 6.9 | 25.0 | 7.2 | 120 | [21] |
| Larval development | 7d-NOEC | - | 2.5 | 4.2 | 2.7 | >99 | ||
| Americamysis bahia | Mortality | 96h-LC50 | 24.0 | 51.0 | 160 | 67.0 | 4100 | [15] |
| Immobility | 96h-EC50 | 19.0 | 48.0 | 92.0 | 47.0 | 4100 | ||
| Mortality | 96h-LC50 | - | - | - | - | 6900 | [9] | |
| Survival | 28d-NOEC | - | - | - | - | 560 | ||
| 28d-LOEC | - | - | - | - | 1100 | |||
| Growth | 28d-NOEC | - | - | - | - | 3900 | ||
| 28d-LOEC | - | - | - | - | >3900 | |||
| Penaeus japonicus | Mortality | 96h-LC50 | 85 | 89 | 71 | 64 | 3900 | [15] |
| Immobility | 96h-EC50 | 31 | 14 | 50 | 20 | 940 | ||
| Crangon uritai | Mortality | 96h-LC50 | 4500 | 360 | 2200 | 1800 | 2200 | [15] |
| Immobility | 96h-EC50 | 3500 | 260 | 570 | 490 | 820 | ||
| Penaeus monodon | Mortality (postlarvae) | 48h-LC50 | >500 | 190 | 408 | - | 390 | [34] |
| Mortality (postlarvae) | 48h-LC10 | - | - | 3 | - | - | [31] | |
| 48h-LC50 | - | - | 175 | - | - | |||
| Callinectes sapidus | Mortality (megalopae) | 24h-LC50 | - | - | 10 | - | - | [35] |
| Mortality (juveniles) | 24h-LC50 | - | - | 1112 | - | - | ||
| Artemia sp. | Mortality | 48h-LC50 | - | - | 361,230 | - | - | [36] |
| Species | Endpoint | Parameter | ACE | CLO | IMI | THI | TMX | Reference |
|---|---|---|---|---|---|---|---|---|
| Daphnia magna | Mortality | 48h-LC50 | - | - | >102,000 | - | >80,000 | [10] |
| Mortality | 48h-LC50 | - | - | 10,440 | - | - | [36] | |
| Ceriodaphnia dubia | Mortality | 48h-LC50 | >33,500 | >100,000 | 72125 | >41,500 | >80,000 | [10] |
| Hyalella azteca | Mortality | 96h-LC50 | 4.8 | 5.2 | 363.2 | 55 | 801 | [10] |
| Immobility | 96h-LC50 | 4.4 | 4.8 | 176.9 | 26.9 | 391 | ||
| Mortality | 7d-LC50 | 4.7 | 4.0 | 230 | 68 | 290 | [37] | |
| Survival | 28d-LC50 | 4.2 | 3.4 | 90 | 44 | 220 | ||
| Growth | 28d-EC50 | 3.4 | 3.5 | 4.3 | 42 | 200 | ||
| Caenis sp. | Mortality | 96h-LC50 | 783 | 122 | <21.8 | 231 | 382 | [10] |
| Immobility | 96h-EC50 | <138.8 | - | <21.8 | <66.3 | <23.3 | ||
| Cloeon sp. | Mortality | 96h-LC50 | 2368 | 3939 | 1152 | 3883 | 4633 | [10] |
| Immobility | 96h-EC50 | <16.6 | <16.4 | 23.1 | 23.1 | 44.1 | ||
| Ephemerella sp. | Mortality | 96h-LC50 | 158.2 | 586.9 | 68.2 | 190.6 | 334.9 | [10] |
| Immobility | 96h-EC50 | <56.1 | 18.5 | 10.6 | <58 | <59 | ||
| Hexagenia sp. | Mortality | 96h-LC50 | >35,600 | >17,400 | 9321 | >9300 | >30,800 | [10] |
| Immobility | 96h-EC50 | 1.8 | 5.5 | n.c. | <1.3 | 35.8 | ||
| Mortality | 96h-LC50 | 780 | 2000 | 900 | 6200 | >10,000 | [37] | |
| Mortality | 96h-NOEC | 1 | 10 | 1 | 1 | 100 | ||
| Behaviour | 96h-EC50 | 4.0 | 24 | 10 | 9.1 | 630 | ||
| Isonychia bicolor | Mortality | 96h-LC50 | >9600 | >1740 | 715 | - | >7120 | [10] |
| Immobility | 96h-EC50 | <600 | <109 | 60.4 | - | <445 | ||
| McCaffertium sp. | Mortality | 96h-LC50 | >890 | 1328 | 1810 | >920 | >920 | [10] |
| Immobility | 96h-EC50 | <56.1 | <109 | 10.6 | 10.6 | 81.7 | ||
| Neocloen triangulifer | Mortality | 96h-LC50 | 1.7 | 3.5 | 5.2 | 1.9 | 5.5 | [10] |
| Immobility | 96h-EC50 | 1.6 | 3.5 | 3.1 | 1.9 | 5.5 | ||
| Coenagrion sp. | Mortality | 96h-LC50 | 24,393 | 14,556 | 3463 | 5647 | 15,062 | [10] |
| Immobility | 96h-EC50 | <5625 | <5919 | <5438 | <2500 | <4188 | ||
| Trichocorixa sp. | Mortality | 48h-LC50 | 1515 | 34.8 | 450.4 | 135.3 | 1473 | [10] |
| Immobility | 48h-EC50 | 63.5 | 21.3 | 63.1 | <39.7 | 56.3 | ||
| Cheumatopsyche sp. | Mortality | 96h-LC50 | 403.8 | 1281 | 324.5 | >920 | 170.1 | [10] |
| Immobility | 96h-EC50 | <56.1 | <108.8 | 176.4 | 162.6 | 118.5 | ||
| Chironomus dilutus | Mortality | 96h-LC50 | 2.8 | 11.6 | 11.8 | 1.6 | 61.9 | [10] |
| Immobility | 96h-EC50 | 2.7 | 3.4 | 2.5 | 0.8 | 36.8 | ||
| Mortality | 14d-LC50 | - | 2.4 | 1.5 | - | 23.6 | [38] | |
| Growth | 14d-EC50 | - | 1.8 | 2.2 | - | 21.4 | ||
| Emergence | 40d-EC50 | - | 0.3 | 0.4 | - | 4.1 | ||
| Mortality | 96h-LC50 | - | - | 7.0 | - | - | [39] | |
| Aedes sp. | Mortality | 48h-LC50 | 159.6 | 28.5 | 40.8 | 53.4 | 67.4 | [10] |
| Aedes aegypti | Mortality | 48h-LC50 | - | - | 44 | - | - | [36] |
| Aedes taeniorhynchus | Mortality | 48h-LC50 | - | - | 13 | - | - | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picone, M.; Distefano, G.G.; Marchetto, D.; Russo, M.; Baccichet, M.; Zangrando, R.; Gambaro, A.; Volpi Ghirardini, A. Inhibition of Larval Development of Marine Copepods Acartia tonsa by Neonicotinoids. Toxics 2022, 10, 158. https://doi.org/10.3390/toxics10040158
Picone M, Distefano GG, Marchetto D, Russo M, Baccichet M, Zangrando R, Gambaro A, Volpi Ghirardini A. Inhibition of Larval Development of Marine Copepods Acartia tonsa by Neonicotinoids. Toxics. 2022; 10(4):158. https://doi.org/10.3390/toxics10040158
Chicago/Turabian StylePicone, Marco, Gabriele Giuseppe Distefano, Davide Marchetto, Martina Russo, Marco Baccichet, Roberta Zangrando, Andrea Gambaro, and Annamaria Volpi Ghirardini. 2022. "Inhibition of Larval Development of Marine Copepods Acartia tonsa by Neonicotinoids" Toxics 10, no. 4: 158. https://doi.org/10.3390/toxics10040158