
The Ecological Role of Ruppia cirrhosa (Petagna) Grande in a Choked Lagoon
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
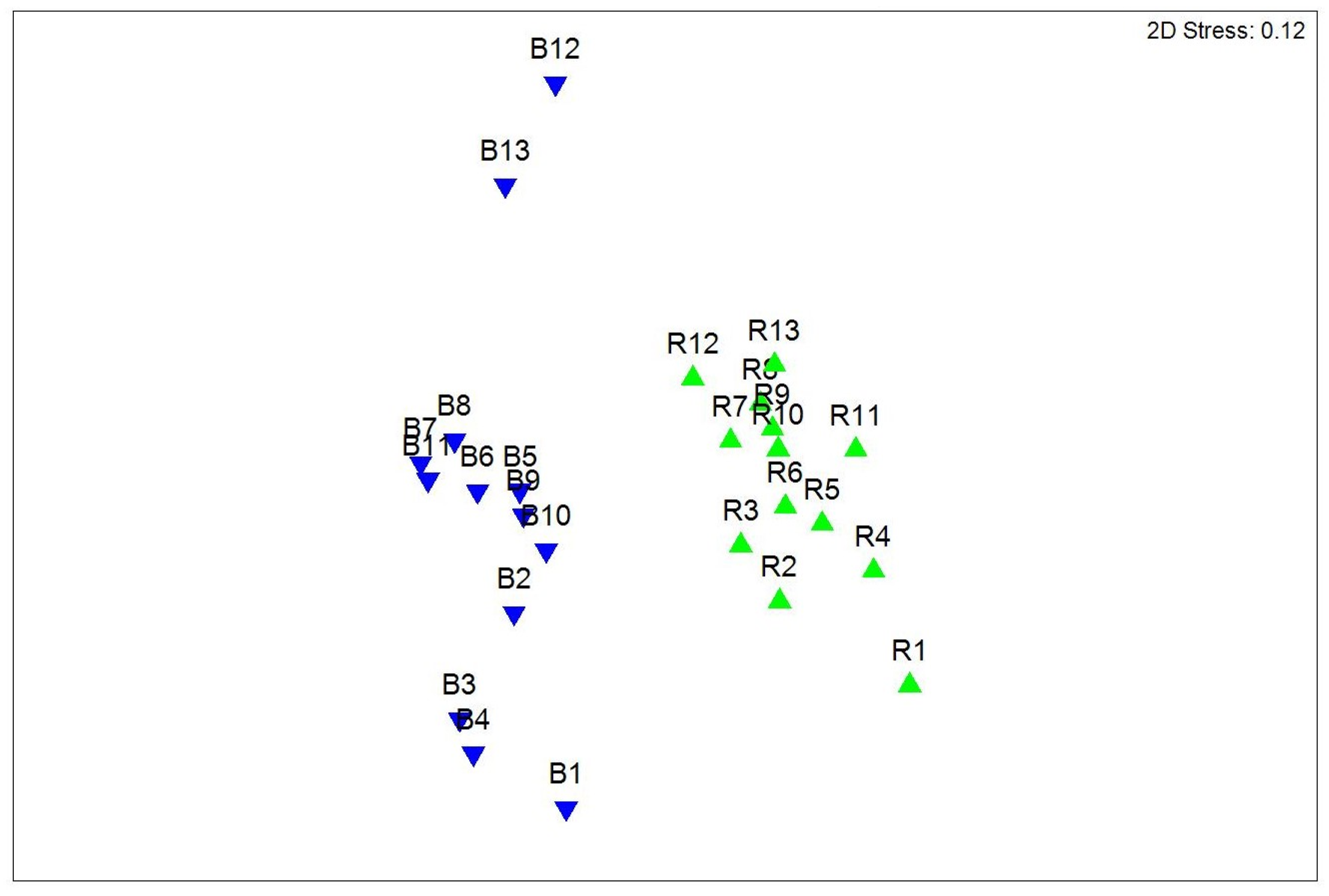
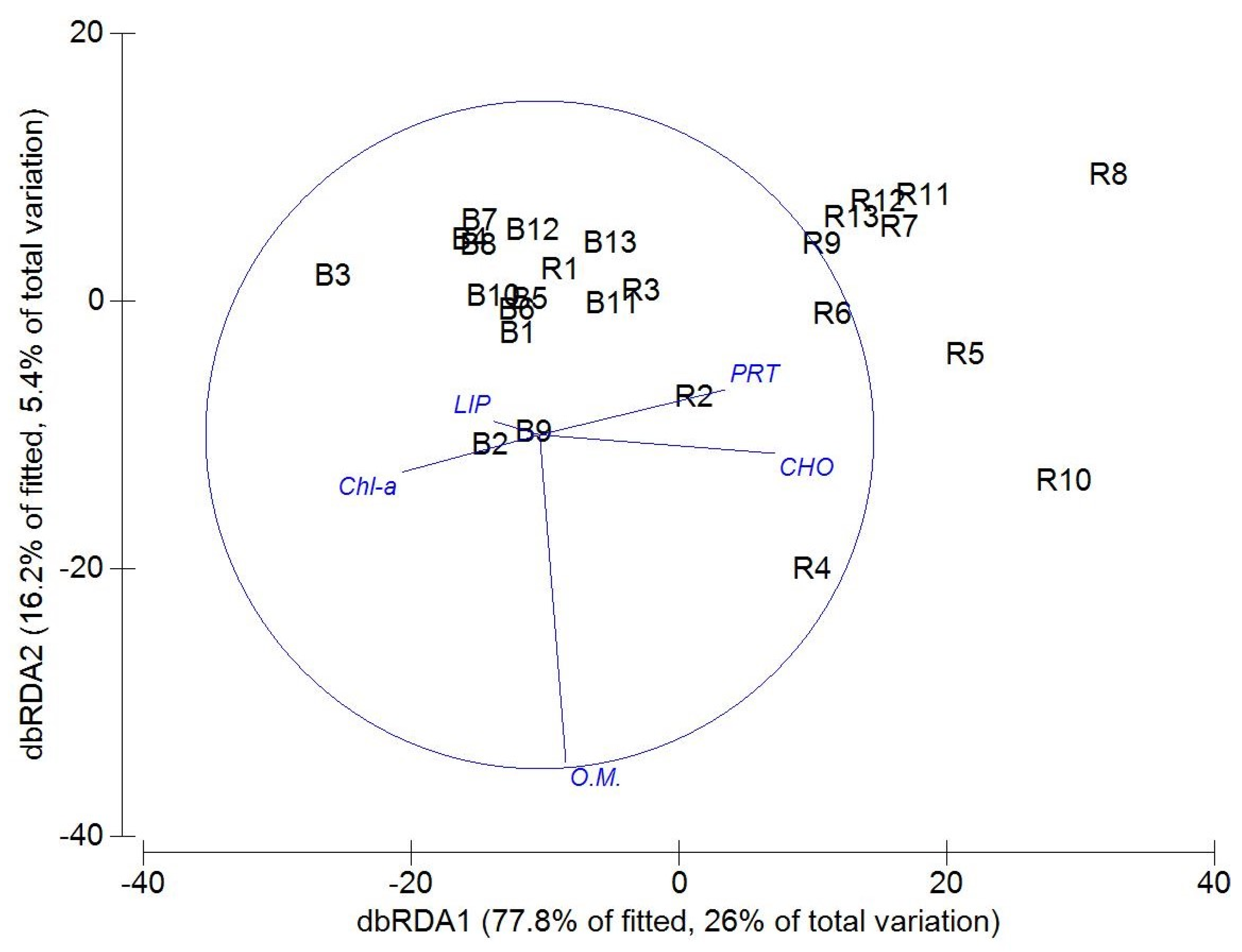
3. Results
3.1. Water and Sediment Parameters
3.2. Macrofaunal Community Biodiversity across a Vegetation Gradient
3.3. Ecological Quality Status
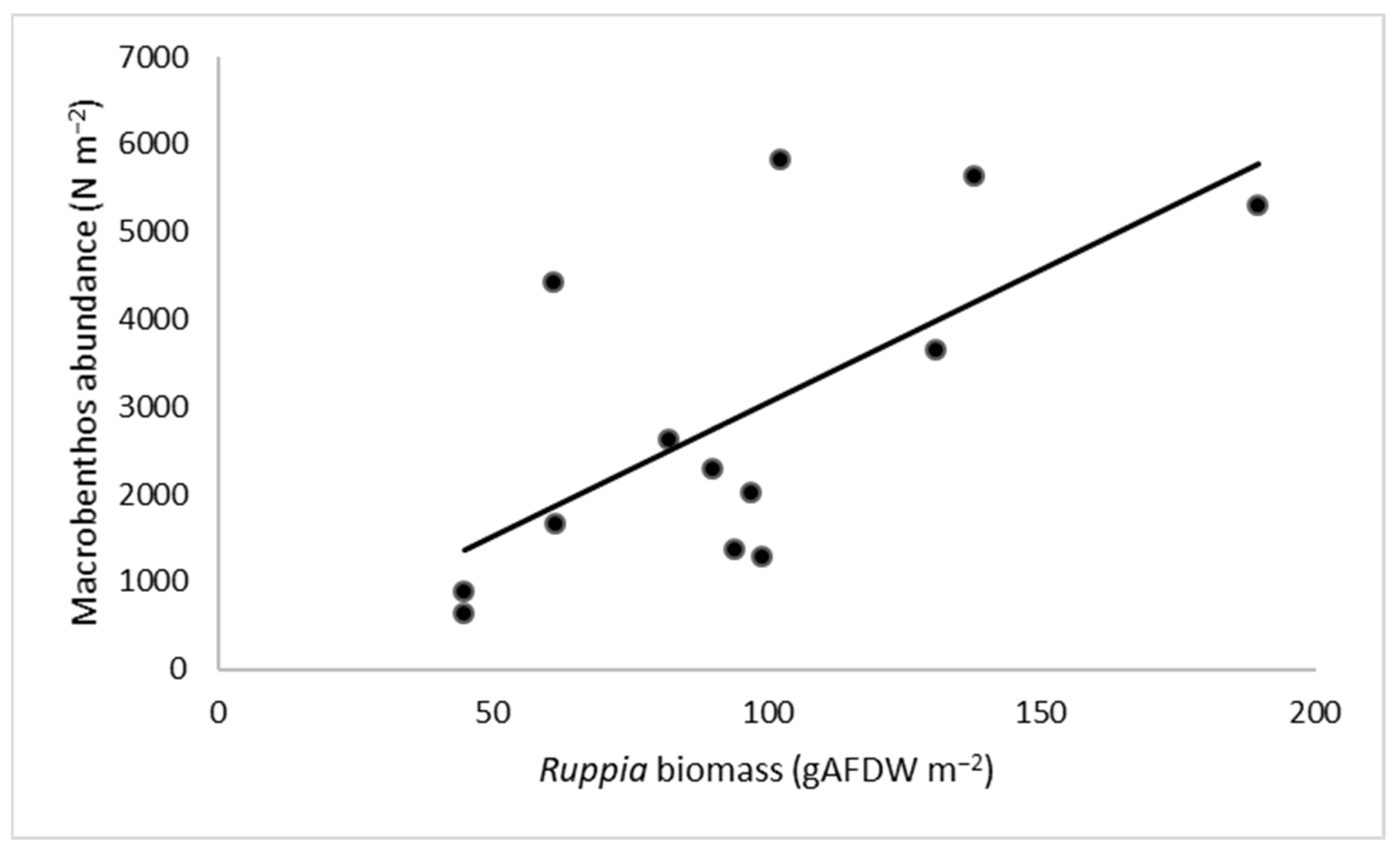
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boström, C.; Jackson, E.L.; Simenstad, C.A. Seagrass landscapes and their effects on associated fauna: A review. Estuar. Coast. Shelf Sci. 2006, 68, 383–403. [Google Scholar] [CrossRef]
- Duarte, C.M.; Marbá, N.; Gacia, E.; Fourqurean, J.W.; Beggins, J.; Barrón, C.; Apostolaki, E.T. Seagrass community metabolism: Assessing the carbon sink capacity of seagrass meadows. Glob. Biogeochem. Cycles 2010, 24, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass ecosystem services and their variability across genera and geographical regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Marcomini, A. Recording the occurrence of trophic level changes in the lagoon of Venice over the ‘90s. Environ. Int. 2005, 31, 993–1001. [Google Scholar]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Abdessalem Shili, A.; Verlaque, M. Regression of Mediterranean seagrasses caused by natural processes and anthropogenic disturbances and stress: A critical review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Boudouresque, C.-F.; Blanfuné, A.; Pergent, G.; Thibaut, T. Restoration of seagrass meadows in the Mediterranean Sea: A critical review of effectiveness and ethical issues. Water 2021, 13, 1034. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Facca, C.; Sfriso, A.A.; Tomio, Y.; Juhmani, A.S.; Wolf, M.A.; Franzoi, P.; Scapin, L.; Pnis, E.; et al. Environmental restoration by aquatic angiosperm transplants in transitional water systems: The Venice Lagoon as a case study. Sci. Total Environ. 2021, 795, 148859. [Google Scholar] [CrossRef]
- Colombo, G. Rassegna delle ricerche condotte nelle Valli da pesca dell’Alto Adriatico. Ann. Università Ferrara 1972, 1, 1–34. [Google Scholar]
- Sorokin, Y.I.; Gnes, A. Structure and functioning of the anthropogenically transformed Comacchio lagoonal ecosystem (Ferrara, Italy). Mar. Ecol. Prog. Ser. 1996, 133, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, J.T.A. The ecology of Ruppia-dominated communities in Western Europe. I. Distribution of Ruppia representatives in relation to their autecology. Aquat. Bot. 1979, 6, 197–268. [Google Scholar] [CrossRef]
- Mannino, A.M.; Menéndez, M.; Obrador, B.; Sfriso, A.; Triest, L. The genus Ruppia L. (Ruppiaceae) in the Mediterranean region: An overview. Aquat. Bot. 2015, 124, 1–9. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A. Synecological classification, structure and dynamics of the macroflora and macrofauna communities. Aquat. Bot. 1980, 8, 1–85. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A. Aspects of production, consumption and decomposition. Aquat. Bot. 1980, 8, 209–253. [Google Scholar] [CrossRef]
- Campagne, C.S.; Salles, J.-M.; Boissery, P.; Deter, J. The seagrass Posidonia oceanica: Ecosystem services identification and economic evaluation of goods and benefits. Mar. Pollut. Bull. 2015, 97, 391–400. [Google Scholar] [CrossRef]
- Sandoval-Gil, J.M.; Marin-Guirao, M.; Ruiz, J.M. The effect of salinity increase on the photosynthesis, growth and survival of the Mediterranean seagrass Cymodocea nodosa. Estuar. Coast. Shelf Sci. 2012, 115, 260–271. [Google Scholar] [CrossRef]
- Triest, L.; Sierens, T. High diversity of Ruppia meadows in saline ponds and lakes of the western Mediterranean. Hydrobiologia 2009, 634, 97–105. [Google Scholar] [CrossRef]
- Lorenzen, C.J. Determination of chlorophyll and phaeopigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Danovaro, R. Methods for the Study of Deep Sea Sediments, Their Functioning and Biodiversity; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015; p. 300. [Google Scholar]
- Muxika, I.; Borja, A.; Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 16–29. [Google Scholar] [CrossRef]
- Bartoli, M.; Nizzoli, D.; Castaldelli, G.; Viaroli, P. Community metabolism and buffering capacity of nitrogen in a Ruppia cirrhosa meadow. J. Exp. Mar. Biol. Ecol. 2008, 360, 21–30. [Google Scholar] [CrossRef]
- Herman, P.M.J.; Middelburg, J.J.; Heip, C.H.R. Benthic community structure and sediment processes on an intertidal flat: Results from the ECOFLAT project. Cont. Shelf Res. 2001, 21, 2055–2071. [Google Scholar] [CrossRef]
- Tolhurst, T.J.; Chapman, M.G.; Murphy, R.J. The Effect of Shading and Nutrient Addition on the Microphytobenthos, Macrofauna, and Biogeochemical Properties of Intertidal Flat Sediments. Front. Mar. Sci. 2020, 7, 419. [Google Scholar] [CrossRef]
- Pusceddu, A.; Sarà, G.; Armeni, M.; Fabiano, M.; Mazzola, A. Seasonal and spatial changes in the sediment organic matter of a semi-enclosed marine system (W-Mediterranean Sea). Hydrobiologia 1999, 397, 59–70. [Google Scholar] [CrossRef]
- Danovaro, R.; Fabiano, M.; Della Croce, N. Labile organic matter and microbial biomasses in deep-sea sediments (Eastern Mediterranean Sea). Deep Sea Res. 1993, 40, 953–965. [Google Scholar] [CrossRef]
- Nelson, W.G. Experimental studies of selective predation on amphipods: Consequences for amphipod distribution and abundance. J. Exp. Mar. Biol. Ecol. 1979, 38, 225–245. [Google Scholar] [CrossRef]
- Henninger, T.O.; Froneman, P.W.; Richoux, N.B.; Hodgson, A.N. The role of macrophytes as a refuge and food source for the estuarine isopod Exosphaeroma hylocoetes (Barnard, 1940). Estuar. Coast. Shelf Sci. 2009, 82, 285–293. [Google Scholar] [CrossRef]
- Boström, C.; Pittman, S.J.; Simentad, C.; Kneib, R.T. Seascape ecology of coastal biogenic habitats: Advances, gaps, and challenges. Mar. Ecol. Prog. Ser. 2011, 427, 191–217. [Google Scholar] [CrossRef] [Green Version]
- Rodil, I.F.; Lohrer, A.M.; Attard, K.M.; Hewitt, J.E.; Thrush, S.F.; Norkko, A. Macrofauna communities across a seascape of seagrass meadows: Environmental drivers, biodiversity patterns and conservation implications. Biodivers. Conserv. 2021, 30, 3023–3043. [Google Scholar] [CrossRef]
- European Commission. Report from the Commission to the European Parliament and the Council on the Implementation of the Water Framework Directive (2000/60/EC) River Basin Management Plans. COM 2012 Brussels, 14.11.2012, 670 Final; European Commission: Brussel, Belgium, 2012. [Google Scholar]
- Pearson, T.H.; Rosenberg, R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev. 1978, 16, 229–311. [Google Scholar]
- Munari, C.; Mistri, M. The performance of benthic indicators of ecological change in Adriatic coastal lagoons: Throwing the baby with the water? Mar. Poll. Bull. 2008, 56, 95–105. [Google Scholar] [CrossRef]
- Dauvin, J.C. Paradox of estuarine quality: Benthic indicators and indices, consensus or debate for the future. Mar. Poll. Bull. 2007, 55, 271–281. [Google Scholar] [CrossRef]
- Munari, C. Effects of the 2003 European heatwave on the benthic community of a severe transitional ecosystem (Comacchio Saltworks, Italy). Mar. Pollut. Bull. 2011, 62, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Christensen, O.B.; Christensen, J.H. Intensification of extreme European summer precipitation in warmer climate. Glob. Planet. Chang. 2004, 44, 107–117. [Google Scholar] [CrossRef]
- Sorokin, Y.I.; Zakuskina, O.Y. Features of the Comacchio ecosystem transformed during persistent bloom of picocyanobacteria. J. Oceanogr. 2010, 66, 373–387. [Google Scholar] [CrossRef]
- Munari, C.; Mistri, M. Ecological status assessment and response of benthic communities to environmental variability: The Valli di Comacchio (Italy) as a study case. Mar. Environ. Res. 2012, 81, 53–61. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 13 June | 30 June | 14 July | 30 July | 10 August | 21 August | 14 September | 22 September | 6 October | 23 October | 8 November | 23 November | 6 December | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ruppia | |||||||||||||
| O.M. | 10.4 | 16.9 | 11.7 | 26.4 | 14.8 | 12.2 | 8.2 | 5.7 | 7.6 | 22.1 | 6.8 | 6.3 | 7.0 |
| Chl-a | 16.15 | 17.40 | 16.92 | 19.08 | 3.11 | 17.73 | 14.29 | 14.06 | 11.66 | 10.74 | 10.94 | 8.19 | 10.00 |
| ±S.E. | 3.40 | 1.89 | 1.13 | 2.98 | 0.22 | 2.82 | 1.99 | 2.47 | 1.21 | 0.56 | 0.32 | 0.40 | 1.07 |
| CHO | 0.66 | 0.91 | 0.80 | 0.83 | 0.98 | 1.24 | 1.23 | 1.07 | 1.15 | 1.28 | 1.16 | 1.08 | 1.10 |
| ±S.E. | 0.05 | 0.17 | 0.04 | 0.12 | 0.13 | 0.05 | 0.18 | 0.05 | 0.09 | 0.07 | 0.06 | 0.07 | 0.10 |
| PRT | 5.65 | 5.91 | 6.40 | 5.50 | 5.83 | 6.38 | 7.43 | 6.41 | 4.57 | 7.69 | 7.58 | 5.90 | 5.63 |
| ±S.E. | 1.60 | 0.73 | 0.32 | 0.22 | 0.64 | 0.09 | 1.33 | 1.91 | 0.24 | 0.32 | 0.02 | 0.56 | 0.16 |
| LIP | 0.94 | 0.99 | 1.07 | 0.72 | 0.97 | 1.06 | 1.24 | 0.63 | 0.76 | 1.28 | 1.26 | 0.98 | 0.94 |
| ±S.E. | 0.27 | 0.12 | 0.05 | 0.04 | 0.11 | 0.01 | 0.22 | 0.32 | 0.04 | 0.05 | 0.00 | 0.09 | 0.03 |
| Bare | |||||||||||||
| O.M. | 13.6 | 18.6 | 10.1 | 8.6 | 11.3 | 12.1 | 8.3 | 8.5 | 17.8 | 11.6 | 12.5 | 8.5 | 8.7 |
| Chl-a | 12.13 | 13.83 | 13.76 | 16.11 | 22.74 | 21.97 | 15.72 | 23.44 | 20.49 | 13.90 | 16.45 | 11.28 | 8.39 |
| ±S.E. | 4.53 | 3.86 | 5.49 | 2.48 | 11.48 | 1.27 | 3.47 | 3.09 | 2.00 | 1.46 | 3.80 | 2.23 | 1.35 |
| CHO | 0.49 | 0.75 | 0.48 | 0.57 | 0.89 | 0.82 | 0.52 | 0.84 | 0.82 | 0.52 | 0.77 | 0.50 | 0.59 |
| ±S.E. | 0.09 | 0.26 | 0.16 | 0.11 | 0.29 | 0.07 | 0.17 | 0.13 | 0.13 | 0.10 | 0.12 | 0.10 | 0.02 |
| PRT | 4.69 | 4.12 | 4.43 | 5.12 | 5.50 | 5.68 | 6.20 | 5.51 | 4.12 | 4.84 | 6.43 | 5.42 | 4.88 |
| ±S.E. | 1.54 | 0.46 | 0.50 | 0.36 | 1.03 | 0.25 | 0.12 | 1.36 | 1.03 | 0.36 | 1.36 | 0.27 | 0.53 |
| LIP | 0.81 | 0.88 | 0.94 | 0.88 | 0.95 | 0.98 | 1.07 | 0.95 | 0.71 | 0.83 | 1.11 | 0.93 | 0.84 |
| ±S.E. | 0.27 | 0.08 | 0.09 | 0.06 | 0.18 | 0.04 | 0.02 | 0.23 | 0.18 | 0.06 | 0.23 | 0.05 | 0.09 |
| Factor | CHL | CHO | |||||
|---|---|---|---|---|---|---|---|
| df | MS | F | p | MS | F | p | |
| Date | 12 | 0.055 | 2.124 | 0.031 | 0.007 | 2.123 | 0.031 |
| Site | 1 | 0.185 | 7.112 | 0.010 | 0.166 | 51.644 | 0.001 |
| Date × Site | 12 | 0.125 | 4.799 | 0.001 | 0.003 | 1.072 | 0.402 |
| Error | 52 | 0.026 | 0.003 |
| Ruppia | Bare | |
|---|---|---|
| Actiniaria sp. | * | |
| Hydrobia acuta | *** | ** |
| Haminaea hydatis | ** | * |
| Cyclope neritea | * | * |
| Littorina obtusata | ** | |
| Bittium reticulatum | * | |
| Cerastroderma glaucum | * | |
| Abra ovata | * | * |
| Flabelligera sp. | * | |
| Hydroides dianthus | * | |
| Alitta succinea | ** | * |
| Capitella capitata | *** | **** |
| Oligochaeta | *** | *** |
| Palaemon elegans | * | |
| Idotea baltica | ** | |
| Gammarus insensibilis | ** | |
| Monocorophium insidiosum | ** | |
| Chironomus salinarus | *** | **** |
| Ecological Groups | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bare | Ruppia | |||||||||
| I (%) | II (%) | III (%) | IV (%) | V (%) | I (%) | II (%) | III (%) | IV (%) | V (%) | |
| 13 June | 0 | 0 | 7.3 | 92.7 | 0 | 5.5 | 2.2 | 41.1 | 51.2 | 0 |
| 30 June | 0 | 0 | 5.3 | 93.7 | 1 | 7.3 | 3 | 9.8 | 73.7 | 6.2 |
| 14 July | 0 | 0 | 8.3 | 87.5 | 4.2 | 0.2 | 4.7 | 8 | 67.9 | 19.2 |
| 30 July | 0 | 0 | 40 | 40 | 20 | 3.9 | 20.8 | 40.6 | 31.9 | 2.8 |
| 10 August | 0 | 0 | 11 | 37.8 | 51.2 | 0.6 | 11.2 | 43.8 | 32.1 | 12.4 |
| 21 August | 0 | 0 | 1.3 | 44.2 | 54.5 | 0.1 | 9.6 | 29.3 | 35.4 | 25.6 |
| 14 September | 0 | 0 | 0 | 72.4 | 27.6 | 0.2 | 0.6 | 9.1 | 40 | 50.2 |
| 22 September | 0 | 0 | 0.6 | 78.9 | 20.5 | 1.5 | 0.9 | 4.1 | 37.5 | 56 |
| 6 October | 0 | 0 | 4.8 | 76.5 | 18.7 | 1.1 | 1.8 | 9.7 | 51.2 | 36.1 |
| 23 October | 0 | 0.7 | 7.3 | 81.7 | 10.2 | 0.3 | 0.7 | 8.4 | 52.7 | 37.9 |
| 8 November | 0 | 0 | 0 | 79.8 | 20.2 | 0.9 | 2.4 | 13.4 | 58.8 | 24.6 |
| 23 November | 1.1 | 0.6 | 0.6 | 0 | 97.8 | 0 | 0.2 | 5.2 | 47.8 | 46.9 |
| 6 December | 0.8 | 0 | 1.6 | 0 | 97.6 | 2.6 | 0.3 | 9.2 | 72.2 | 15.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munari, C.; Casoni, E.; Cozzula, C.; Pasculli, A.; Pezzi, M.; Sciuto, K.; Sfriso, A.A.; Sfriso, A.; Mistri, M. The Ecological Role of Ruppia cirrhosa (Petagna) Grande in a Choked Lagoon. Water 2023, 15, 2162. https://doi.org/10.3390/w15122162
Munari C, Casoni E, Cozzula C, Pasculli A, Pezzi M, Sciuto K, Sfriso AA, Sfriso A, Mistri M. The Ecological Role of Ruppia cirrhosa (Petagna) Grande in a Choked Lagoon. Water. 2023; 15(12):2162. https://doi.org/10.3390/w15122162
Chicago/Turabian StyleMunari, Cristina, Elia Casoni, Cinzia Cozzula, Alessandra Pasculli, Marco Pezzi, Katia Sciuto, Andrea Augusto Sfriso, Adriano Sfriso, and Michele Mistri. 2023. "The Ecological Role of Ruppia cirrhosa (Petagna) Grande in a Choked Lagoon" Water 15, no. 12: 2162. https://doi.org/10.3390/w15122162